


Research on Improving the Thermostability of Phytase YiAPPA by Introducing Ionic Bonds through Site-directed Mutagenesis
-
摘要: 本研究旨在通过定点突变引入离子键来提高植酸酶YiAPPA的热稳定性。对比YiAPPA与热稳定性优良的植酸酶rPhyXT52的分子结构,采用定点突变技术向YiAPPA中引入与rPhyXT52热稳定性相关的分子表面离子键,构建离子键突变体。通过热稳定性筛选,获得热稳定性显著提高的离子键突变体T209K/S220E/N237D。突变体T209K/S220E/N237D的酶学特性研究结果表明:突变体T209K/S220E/N237D于37 ℃、pH4.5的绝对酶活为3982.06 U/mg,与YiAPPA基本一致;T209K/S220E/N237D的最适反应温度、最适反应pH、pH稳定性以及蛋白酶抗性也与YiAPPA基本一致;与YiAPPA相比,T209K/S220E/N237D于80 ℃半衰期由14.81 min延长至24.72 min,半失活温度T5030由55.12 ℃提升至64.05 ℃,Tm值由48.36 ℃提升至55.04 ℃。分子动力学模拟显示,T209K/S220E/N237D中引入了新的离子键,提高了酶分子中构成这些离子键的氨基酸残基所在的结构单元的稳定性,从而提高了酶的热稳定性。本研究结果表明,向YiAPPA中引入离子键可有效提高其热稳定性,使其更适用于食品加工领域。本研究也可为植酸酶以及其他类型酶的热稳定性改造提供理论依据。Abstract: To improve the thermostability of the phytase YiAPPA by introducing ionic bonds through site-directed mutagenesis. By comparing the molecular structure of YiAPPA and phytase rPhyXT52 with excellent thermal stability, site-directed mutagenesis was used to introduce molecular surface ionic bonds related to the thermostability of rPhyXT52 into YiAPPA to construct mutants. The mutant T209K/S220E/N237D with significantly improved thermo stability were obtained through screening. The mutant T209K/S220E/N237D had an specific activity of 3982.06 U/mg at 37 °C and pH4.5, which was essentially consistent with the respective values for YiAPPA. The optimal reaction temperature, optimal reaction pH, pH stability, and protease resistance of T209K/S220E/N237D were also basically the same as those of YiAPPA. However, compared with that of YiAPPA, the half-life of T209K/S220E/N237D at 80 °C was extended from 14.81 to 24.72 min, the half-inactivation temperature T5030 was increased from 55.12 to 64.05 °C, and the Tm value was increased from 48.36 ℃ to 55.04 ℃. Molecular dynamics simulations showed that the introduction of new ionic bonds in T209K/S220E/N237D improved the stability of the structural units in the enzyme molecule in which the amino acid residues constituting these ionic bonds were located, thereby improving the thermostability of the enzyme. Results showed that the introduction of ionic bonds into YiAPPA could effectively improve its thermostability and make it more applicable for food processing. The results would be also provide a theoretical basis for the improved thermostability of phytase and other enzymes.
-
Keywords:
- phytase /
- site-directed mutagenesis /
- ionic bonds /
- thermostability /
- molecular dynamics simulation
-
植酸是豆类、谷类等作物中磷的主要存在形式,植酸在生物体内与金属阳离子、氨基酸、蛋白质和淀粉等物质结合从而抑制这些物质的消化吸收[1−2]。因此,植酸通常被认为是抗营养因子。植酸酶又称肌醇六磷酸酶,可以降解植酸生成无机磷和磷酸肌醇,从而解除植酸的抗营养作用[3−4]。人体消化系统缺乏植酸酶,因此不能代谢植酸,进而限制了营养素在肠道的消化吸收,降低了食品的营养价值。已有研究表明,相较于烹饪、热水煮、发酵、浸泡等食品加工方法,使用植酸酶可更有效地减少食品中植酸含量[5−8]。在植物性食品原料中添加植酸酶可降解植酸盐释放无机磷,一方面可以提高食品中磷的吸收利用率;另一方面有利于解除植酸的抗营养作用。因此植酸酶可以作为一种新型食品添加剂应用于食品工业。然而,食品工业的许多工艺过程需要在高温条件下进行,天然植酸酶热稳定性差是限制植酸酶在食品工业应用的关键问题[9−10]。因此寻找和开发热稳定性优良的植酸酶仍是目前国内外学者研究的热点。
理性设计方法基于酶分子结构与功能之间的构效关系对酶进行定向改造,理性设计方法包括离子键设计、二硫键设计、氢键设计、糖基化位点设计、同源模建、分子动力学模拟等,理性设计已被广泛应用于提高植酸酶热稳定性[11−13]。例如,Liao等[14]向来源于Aspergillus niger的植酸酶PhyA中引入离子键,构建热稳定性提高的突变体I44E/T252R,其于80 ℃保温10 min后残留酶活力提高了20%。Zhang等[15]向来源于Bacillus licheniformis WHU的植酸酶中引入二硫键,获得了60 ℃半衰期提高了3.8倍的突变体G197C/A358C。Wang等[16]向大肠杆菌植酸酶中引入氢键,获得了Tm值提高3.8 ℃的突变体K74D/K75Q/K180N/R181N/K183S/K363N。Li等[17]基于来源于Escherichia coli的植酸酶的分子结构,通过氨基酸残基温度因子(B-factor)分析软件B-FITTER计算植酸酶中各氨基酸残基的B-factor,确定其中13个氨基酸残基作为突变位点,构建突变体文库并筛选出热稳定性提高的突变体P56214,使其于90 ℃保温5 min后的残留酶活由20%提高至75%。
来源于Yersinia intermedia的植酸酶YiAPPA(EC 3.1.3.2)是目前已知的植酸酶活性最高的植酸酶,其酶活高达3960 U/mg(37 ℃、pH4.5)[18−19]。YiAPPA具有酸性条件下的高稳定性和高酶活、较强的蛋白酶抗性等突出性质,因此其在食品加工领域具有巨大的应用潜力。但是YiAPPA的热稳定性(YiAPPA于80 ℃的半衰期仅约为15 min)制约了其在食品加工领域的应用。由宏基因组筛选得到的植酸酶rPhyXT52具有优良的热稳定性(于80 ℃的半衰期约为2.1 h),其与YiAPPA的序列相似度为48.43%,并且已有研究证实rPhyXT52分子表面离子键对于维持其热稳定性非常重要[20]。为了提高YiAPPA的热稳定性,本研究拟将rPhyXT52中离子键引入至YiAPPA中,以期获得热稳定性提高的突变体,并通过分子动力学模拟分析,探究突变体热稳定性提高的分子机制,为植酸酶及其他类型酶的热稳定性改造提供理论依据。
1. 材料与方法
1.1 材料与仪器
大肠杆菌Escherichia coli JM109、枯草芽孢杆菌Bacillus subtilis WB600、重组质粒pSTOP1622-yiappah 均由本实验室保存;KOD-Plus-neo DNA聚合酶 日本Toyobo公司限制性内切酶、DNA Marker、蛋白质Marker、质粒DNA小量纯化试剂盒 日本TaKaRa公司;QuickMutation™基因定点突变试剂盒 上海碧云天生物技术有限公司;Chelating SepharoseTM Fast Flow 美国GE Healthcare公司;Bradford法蛋白质定量检测试剂盒 上海生工生物工程股份有限公司;实验所用试剂均为国产分析纯。
Mastercycler gradient型PCR仪 美国Eppendorf公司;TY04S-3C型凝胶成像系统 北京君意东方电泳设备有限公司;SP-752PC型紫外可见分光光度计 上海光谱仪器有限公司; Nano-DSC型微量差示扫描热量仪 美国TA仪器公司;iMark 酶标仪 美国Bio-Rad公司。
1.2 实验方法
1.2.1 生物信息学分析
以来源于Y. kristensenii的植酸酶YkAPPA的分子结构(PDB ID:4ARV)为模板[21−22],采用SWISS-MODEL(http://swissmodel.expasy.org)[23]分别同源模建植酸酶YiAPPA和rPhyXT52的蛋白质分子结构。采用三维图像软件PyMOL v0.99 显示蛋白质分子结构。将植酸酶的蛋白质分子结构提交至在线工具RING-2.0 (http://protein.bio.unipd.it/ring)[24],分析蛋白质分子表面离子键的分布情况。
1.2.2 离子键突变体的构建与鉴定
根据QuickMutation™基因定点突变试剂盒的说明书,结合基因yiappa的碱基序列和拟突变的氨基酸位点设计引物,如表1所示。以定点突变体T47E/Q50K的构建为例,以重组质粒pSTOP1622-yiappah为模板,采用引物T47E/Q50K-F和T47E/Q50K-R,经PCR扩增得到包含载体序列和突变基因序列的线性片段。PCR扩增条件为:95 ℃ 5 min;95 ℃ 30 sec,58 ℃ 30 sec,68 ℃ 3 min,35个循环;68 ℃,10 min。PCR扩增产物经限制性内切酶DpnⅠ处理后,直接转化E. coli JM109,涂布于卡那霉素抗性平板进行筛选。从卡那霉素抗性平板挑取转化子,提取重组质粒,将重组质粒送至上海生工生物工程股份有限公司进行测序,并与相应基因序列进行比对确认。其他定点突变体的构建方法参照突变体T47E/Q50K的构建方法。
表 1 构建重组质粒所用引物Table 1. Primers for constructing recombinant plasmids引物名称 序列 T47E/Q50K-F 5’-GTTACCCCTGACGAGTGGCCGAAATGGC
CGGTCGCCGCAG-3’T47E/Q50K-R 5’-CTGCGGCGACCGGCCATTTCGGCCACTCGT
CAGGGGTAAC-3’Q64E-F 5’-CGAGGTGCAGAATTAGTGACATTGATGG-3’ Q64E-R 5’-CCATCAATGTCACTAATTCTGCACCTCG-3’ T148D-F 5’-GTAAGTTAGATTCGGATCAAACCCATAAGGC-3’ T148D-R 5’-GCCTTATGGGTTTGATCCGAATCTAACTTAC-3’ N181D-F 5’-GATGGGTGAGATTCTGGATTTTGCGGCATCTC-3’ N181D-R 5’-GAGATGCCGCAAAATCCAGAATCTCACCCATC-3’ S220E-F 5’-CAAAAGTCTCGCTCGAAGGACCACTGGCACTG-3’ S220E-R 5’-CAGTGCCAGTGGTCCTTCGAGCGAGACTTTTG-3’ T209K-F 5’-CAGCGAATAAGATCAAGGTGAACAAGCCG-3’ T209K-R 5’-CGGCTTGTTCACCTTGATCTTATTCGCTG-3’ N237D-F 5’-GATCTTTTTGCTACAAGATTCACAAGCGATGC-3’ N237D-R 5’-GCATCGCTTGTGAATCTTGTAGCAAAAAGATC-3’ 注:方框标注的部分为突变的碱基。 1.2.3 重组植酸酶的诱导表达与纯化
枯草芽孢杆菌B. subtilis WB600感受态细胞的制备和转化采用改进的Spizizen 法进行[25]。将重组质粒转入枯草芽孢杆菌B. subtilis WB600感受态细胞,获得重组枯草芽孢杆菌。重组植酸酶的诱导表达与纯化参照文献[19]进行。采用聚丙烯酰胺凝胶电泳(Sodium dodecyl sulphate-polyacrylamide gel electrophoresis,SDS-PAGE) [26]检测重组植酸酶,根据蛋白质样品在SDS-PAGE图谱上所呈现的带型特点来判断重组植酸酶的纯度,并采用Bradford法[27]测定重组植酸酶的浓度。
1.2.4 重组植酸酶的酶活力测定
将250 μL酶液(0.005 mg/mL)与750 μL 0.25 mol/L醋酸钠缓冲液(pH 4.5)混匀后加入离心管中,作为重组植酸酶的酶活力测定样品。准备两份样品,分别设立为实验组和对照组。向实验组中加入2 mL 1.5 mmol/L 植酸钠溶液(0.25 mol/L醋酸钠缓冲液,pH4.5),向对照组中加入2 mL终点混合液(钼酸铵/钒酸铵/硝酸),充分混匀。于37 ℃反应30 min后,立即向实验组中加入2 mL终点混合液,对照组中加入2 mL 1.5 mmol/L植酸钠溶液,并充分混匀,然后于415 nm处测定光吸收值。植酸酶活力单位(U)定义为:在37 ℃、pH4.5的条件下,每分钟从1.5 mmol/L植酸钠溶液中释放出1 μmol/L无机磷所需要的植酸酶量为一个酶活力单位(U)。
1.2.5 重组植酸酶的酶学性质测定
重组植酸酶的最适反应温度、热稳定性、最适反应pH、不同pH下稳定性、蛋白酶抗性测定均参照文献[19]进行。
根据“1.2.4”配制反应体系(即向实验组中加入2 mL 1.5 mmol/L 植酸钠溶液,向对照组中加入2 mL终点混合液,充分混匀),并将反应体系分别于30~90 ℃反应30 min,测定不同温度条件下样品的酶活力,将最高酶活力定义为100%,计算各个温度下的相对酶活,并以相对酶活对温度作图,确定其最适反应温度。将酶液于80 ℃保温0~40 min或100 ℃保温0~20 min,根据“1.2.4”测定样品的酶活力,将未处理样品的酶活力定义为100%,计算其余样品的残留酶活力,以残留酶活力对温度作图,评价酶于80 ℃或100 ℃的稳定性。
将250 μL酶液(0.005 mg/mL)与750 μL 不同pH(1.0~8.0)缓冲液混匀后加入离心管中,根据“1.2.4”测定样品的酶活力,将最高酶活力定义为100%,计算各pH下的相对酶活,并以相对酶活对pH作图,确定其最适反应pH。将250 μL酶液(0.005 mg/mL)与750 μL 不同pH(1.0~12.0)缓冲液混匀后加入离心管中,于37 ℃处理2 h,然后用最适pH缓冲液将样品稀释4倍,再测定稀释后样品的酶活力,以未经37 ℃处理2 h的样品的酶活力定义为100%,计算保温处理后样品的残留酶活力,以残留酶活力对pH作图,评价酶于不同pH下稳定性。所使用的缓冲液如下:0.25 mol/L 甘氨酸-盐酸缓冲液,pH1.0~3.5;0.25 mol/L 醋酸钠-醋酸缓冲液,pH3.5~6.0;0.25 mol/L Tris-盐酸缓冲液,pH6.0~8.5;0.25 mol/L 甘氨酸-氢氧化钠缓冲液,pH8.5~12.0。
分别配制0.1 mg/mL的胃蛋白酶(0.25 mol/L Gly-HCl缓冲液,pH2.0)和胰蛋白酶(0.25 mol/L Tris-HCl缓冲液,pH7.0),按照蛋白酶与植酸酶质量比为1:10的比例向植酸酶中分别加入胃蛋白酶和胰蛋白酶,于37 ℃分别保温2 h后,向其中加入蛋白酶抑制剂终止反应,然后用最适pH缓冲液(0.25 mol/L醋酸钠缓冲液,pH4.5)对蛋白酶处理的样品进行100倍稀释,再根据“1.2.4”测定植酸酶处理前后的酶活力。
1.2.6 重组植酸酶的热力学参数测定
重组植酸酶的热力学参数表征主要包括半衰期(t1/2)、半失活温度(T5030)和热解折叠温度(Tm)。
将酶液(0.005 mg/mL)于80 ℃保温,测定不同时间下的酶活力。以未处理样品的酶活力为100%,计算其余样品的残留酶活力。以残留酶活力的对数为纵坐标,以时间t为横坐标,进行线性拟合,计算重组植酸酶的失活动力学常数kd。利用一级失活方程(1)[28−29]计算重组植酸酶的半衰期(t1/2)。一级失活方程中A0为初始酶活力,At为不同时间下酶活力。
At=A0e−kd×t (1) 将酶液(0.005 mg/mL)分别于45、50、55、60、65、70 ℃下保温30 min,测定保温后样品的酶活力。以未处理样品的酶活力为100%,计算不同温度下保温处理后样品的残留酶活力。残留酶活力为50%时所对应的温度即为半失活温度(T5030)。
采用Nano-DSC型微量差示扫描热量仪测定重组植酸酶的热解折叠温度(Tm)。首先将样品脱气、上样,待热流信号和压力信号稳定后,开始以1 ℃/min的速率使温度从30 ℃上升至80 ℃对样品进行扫描。利用软件Nano Analysis对测试结构进行拟合,得出热解折叠温度(Tm)。
1.2.7 重组植酸酶的动力学常数测定
用0.25 mol/L醋酸钠缓冲液(pH4.5)配制不同浓度的植酸钠溶液(0.0625%、0.1%、0.125%、0.2%、0.25%、0.5%、1.0%、1.5%),向不同浓度的植酸钠溶液中加入等量的酶液,按照“1.2.4”测定酶活力。根据双倒数作图法[30]以底物浓度([S])的倒数为横坐标,以酶活力(V)的倒数为纵坐标作图,直线的斜率为Km/Vmax,截距为1/Vmax,计算以植酸钠为底物时的米氏常数Km、最大反应速度Vmax。根据式(3)计算反应常kcat,其中[Et]为酶液的浓度。
1V=KmVmax×1[S]+1Vmax (2) kcat=Vmax[Et] (3) 1.2.8 分子动力学模拟
采用程序Gromacs 2019.6[31]在温度373 K进行100 ns的分子动力学模拟,分析重组植酸酶的热力学波动性。分子动力学模拟在恒温恒压以及周期性边界条件下进行,应用Amber14SB全原子力场,TIP3P水模型。运行结束后即可获得反映酶整体分子结构刚性的RMSD文件和反映酶分子中氨基酸残基波动性大小的RMSF文件。
1.3 数据处理
所有试验均重复3次,试验结果用平均值±标准差表示,运用软件SigmaPlot 14.0对试验数据进行统计分析并作图。
2. 结果与分析
2.1 离子键突变体的构建与初步筛选
2.1.1 离子键突变体的构建
据文献文献[20]报道,植酸酶rPhyXT52于80 ℃的半衰期约为2.1 h,植酸酶YiAPPA于80 ℃的半衰期约为15 min[18],即rPhyXT52的热稳定性明显优于YiAPPA,并且Tan等[20]通过比较rPhyXT52及其分子表面离子键缺失突变体的热稳定性,证实rPhyXT52分子表面离子键对于维持热稳定性非常重要。rPhyXT52和YiAPPA同属于组氨酸酸性磷酸酶,具有相同的催化机制和相似的分子结构。本研究对rPhyXT52和YiAPPA分子表面离子键进行分析,将rPhyXT52分子表面离子键引入至YiAPPA中,以期获得热稳定性提高的YiAPPA突变体。
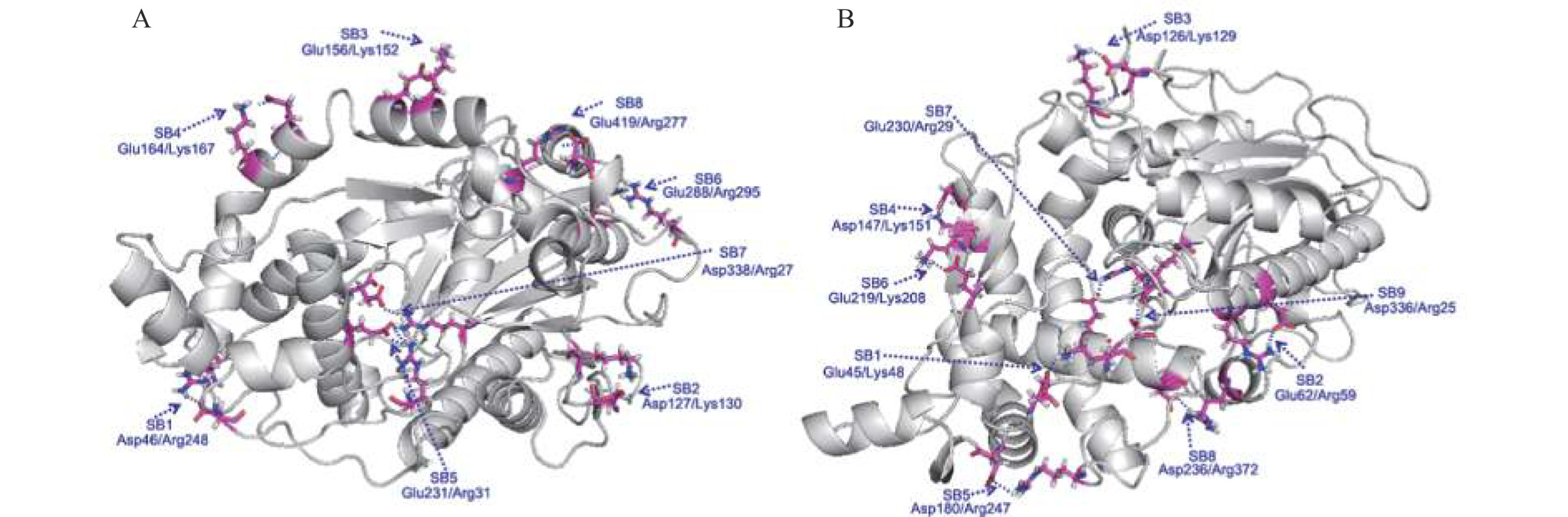
将植酸酶YiAPPA和rPhyXT52的蛋白质序列提交至在线网站SWISS-MODEL (http://swissmodel.expasy.org),同源模建两种植酸酶的蛋白质分子结构。进一步将两种植酸酶的蛋白质分子结构提交至在线工具RING-2.0(http://protein.bio.unipd.it/ring),分析其分子表面离子键的分布情况。同源模建结果显示,植酸酶YiAPPA和rPhyXT52的蛋白质分子结构均以植酸酶YkAPPA的分子结构(PDB ID:4ARV)为模板同源模建得到。植酸酶YiAPPA和rPhyXT52的蛋白质分子结构以及分子表面离子键的分布情况如图1所示,两种植酸酶的分子表面离子键对比分析结果如表2所示。植酸酶YiAPPA有8对分子表面离子键,植酸酶rPhyXT52有9对分子表面离子键。通过YiAPPA与rPhyXT52的蛋白质序列以及三维分子结构比对发现,rPhyXT52中构成其中3对分子表面离子键的氨基酸残基(Asp126-Lys129、Glu230-Arg29、Asp336-Arg25)与YiAPPA中构成其中3对分子表面离子键的氨基酸残基(Asp127-Lys130、Glu231-Arg31、Asp338-Arg27)相互对应,即YiAPPA与rPhyXT52中有3对分子表面离子键是相互对应的。
![]() 图 1 植酸酶YiAPPA(A)和植酸酶rPhyXT52(B)分子表面离子键注:图中蓝色虚线标记的均为氨基酸残基对之间形成的离子键。Figure 1. Surficial salt bridges of phytase YiAPPA (A) and phytase rPhyXT52 (B)表 2 植酸酶rPhyXT52分子表面离子键与植酸酶YiAPPA分子表面离子键对比分析Table 2. Comparative analysis of surficial salt bridges between phytase rPhyXT52 and phytase YiAPPA
图 1 植酸酶YiAPPA(A)和植酸酶rPhyXT52(B)分子表面离子键注:图中蓝色虚线标记的均为氨基酸残基对之间形成的离子键。Figure 1. Surficial salt bridges of phytase YiAPPA (A) and phytase rPhyXT52 (B)表 2 植酸酶rPhyXT52分子表面离子键与植酸酶YiAPPA分子表面离子键对比分析Table 2. Comparative analysis of surficial salt bridges between phytase rPhyXT52 and phytase YiAPPArPhyXT52分子
表面
离子键rPhyXT52分子表面离子键及相关
氨基酸残基
在YiAPPA中对应情况YiAPPA分子
表面
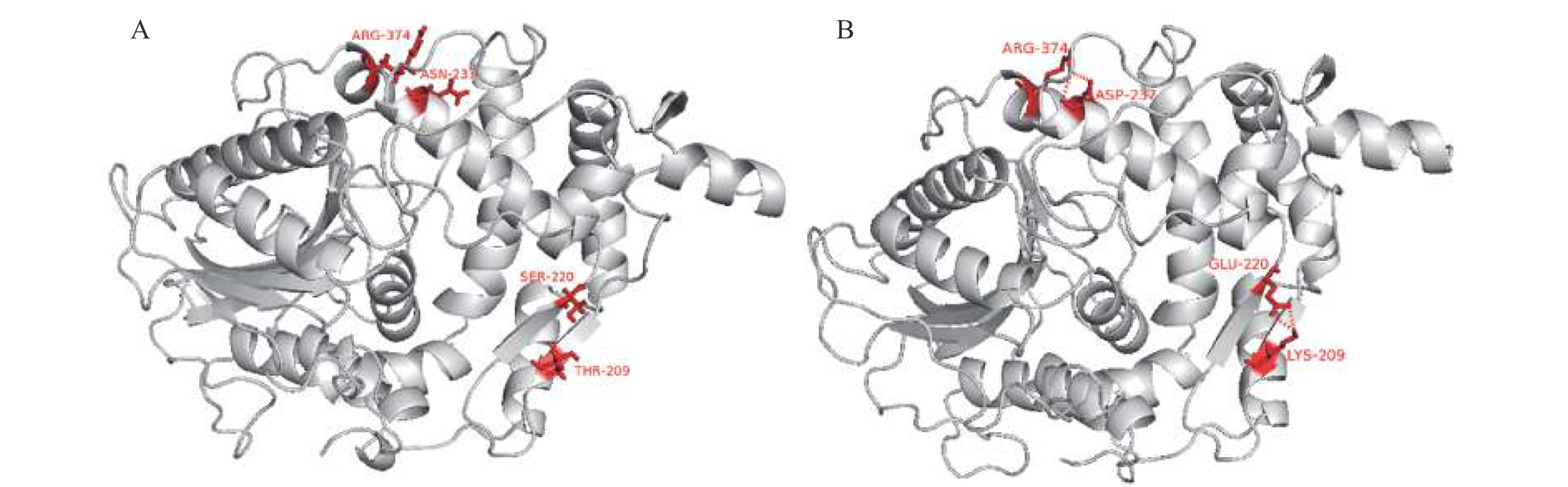
离子键Glu45-Lys48 无对应离子键,Thr47-Gln50 Asp46-Arg248 Glu62-Arg59 无对应离子键,Gln64-Arg61 Asp127-Lys130 Asp126-Lys129 有对应离子键,Asp127-Lys130 Glu156-Lys152 Asp147-Lys151 无对应离子键,Thr148-Lys152 Glu164-Lys167 Asp180-Arg247 无对应离子键,Gln181-Arg248 Glu231-Arg31 Glu219-Lys208 无对应离子键,Ser220-Thr209 Glu288-Arg295 Glu230-Arg29 有对应离子键,Glu231-Arg31 Asp338-Arg27 Asp236-Arg372 无对应离子键,Asn237-Arg374 Glu419-Arg277 Asp336-Arg25 有对应离子键,Asp338-Arg27 注:下划线标注的是YiAPPA中拟进行定点突变的氨基酸残基。 基于以上分析,本研究拟将rPhyXT52中其余6对分子表面离子键引入至YiAPPA中,即将如下氨基酸残基突变引入至YiAPPA中:T47E/Q50K、Q64E、T148D、N181D、T209K/S220E、N237D。其中氨基酸残基突变T47E/Q50K可能引入新的离子键E47-K50;氨基酸残基突变Q64E可能引入新的离子键E64-R61;氨基酸残基突变T148D可能引入新的离子键D148-K152;氨基酸残基突变N181D可能引入新的离子键D181-R248;氨基酸残基突变T209K/S220E可能引入新的离子键K209-E220;氨基酸残基突变N237D可能引入新的离子键D237-R374。采用SWISS-MODEL同源模建包含以上6个氨基酸残基突变的突变体T47E/Q50K/Q64E/T148D/N181D/T209K/S220E/N237D的蛋白质分子结构,并采用在线工具RING-2.0分析突变体表面离子键,结果显示(图2):仅氨基酸突变T209K/S220E及N237D引入了新的离子键,其余氨基酸残基突变未引入新的离子键(结果未显示)。因此,本研究构建了离子键突变体T209K/S220E、N237D以及T209K/S220E/N237D,并比较了突变体与YiAPPA的酶学性质。
![]() 图 2 植酸酶的分子结构分析注:A:植酸酶YiAPPA的分子结构分析;B:突变体T47E/Q50K/Q64E/T148D/N181D/T209K/S220E/N237D的分子结构分析;图中红色标记的氨基酸残基为突变的氨基酸残基以及与其形成新的离子键的氨基酸残基(Lys209与Glu220形成新的离子键;Asn237与Arg374形成新的离子键);红色虚线标记的为引入的离子键。Figure 2. Molecular structure analysis of recombinant phytase
图 2 植酸酶的分子结构分析注:A:植酸酶YiAPPA的分子结构分析;B:突变体T47E/Q50K/Q64E/T148D/N181D/T209K/S220E/N237D的分子结构分析;图中红色标记的氨基酸残基为突变的氨基酸残基以及与其形成新的离子键的氨基酸残基(Lys209与Glu220形成新的离子键;Asn237与Arg374形成新的离子键);红色虚线标记的为引入的离子键。Figure 2. Molecular structure analysis of recombinant phytase2.1.2 离子键突变体的初步筛选
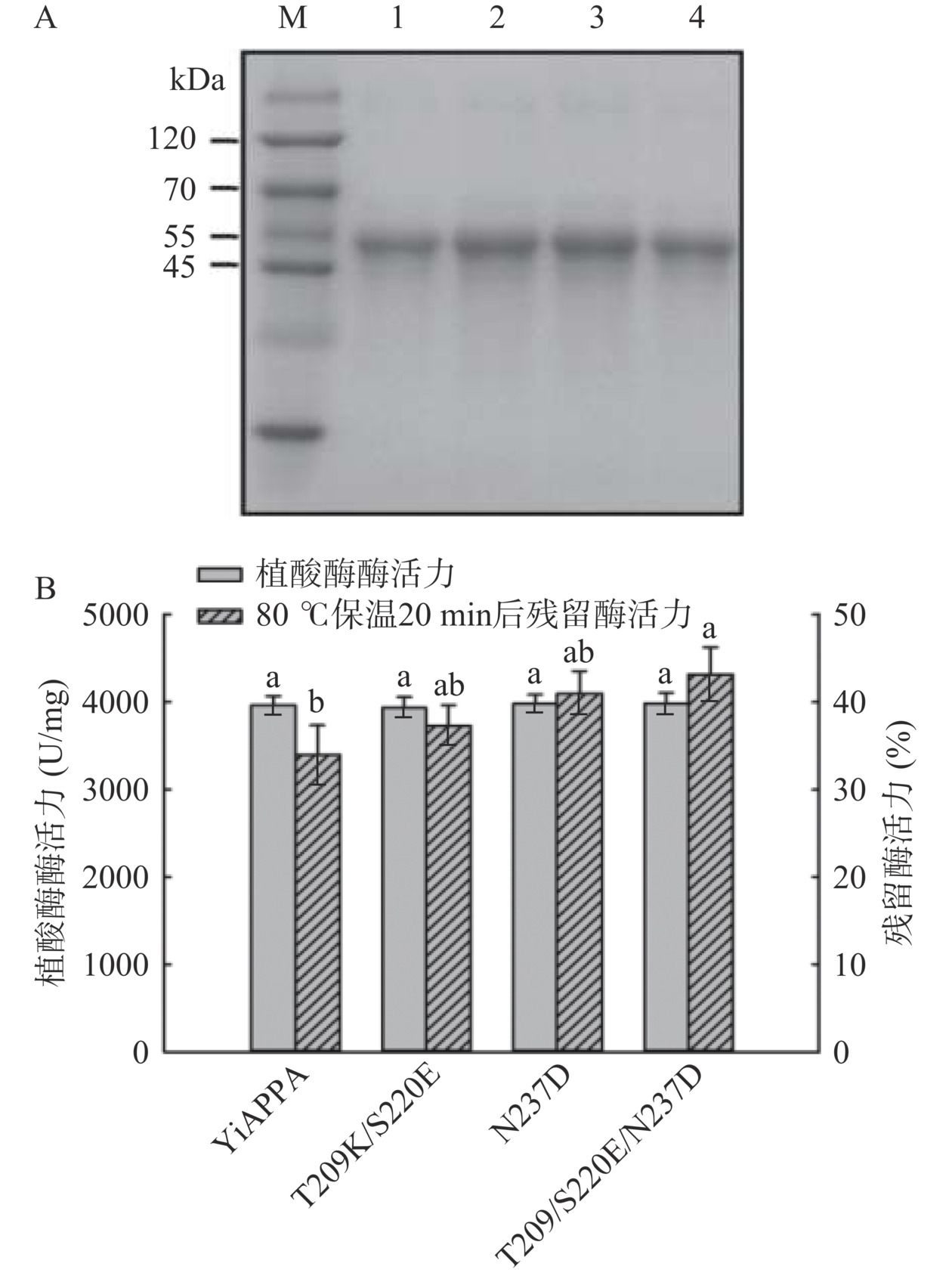
将YiAPPA及其离子键突变体T209K/S220E、N237D以及T209K/S220E/N237D分别于枯草芽孢杆菌B. subtilis WB600中进行异源重组表达,并对表达得到的重组植酸酶进行分离纯化,采用SDS-PAGE检测重组植酸酶的纯度,同时测定重组植酸酶的绝对酶活力及其于80℃保温20 min的残留酶活力。如图3A所示,YiAPPA及其离子键突变体均得到重组表达并纯化为单一条带,SDS-PAGE检测结果显示YiAPPA及其离子键突变体的分子量均约为48 kDa,与野生型YiAPPA的分子量基本一致[18]。图3B显示重组YiAPPA的绝对酶活为3960.81 U/mg;突变体T209K/S220E的绝对酶活为3935.18 U/mg,突变体N237D的绝对酶活为3977.23 U/mg,突变体T209K/S220E/N237D的绝对酶活为3982.06 U/mg,与重组YiAPPA基本一致。重组YiAPPA于80 ℃保温20 min后的残留酶活力为33.99%;突变体T209K/S220E、N237D及T209K/S220E/N237D于80 ℃保温20 min后的残留酶活力均明显高于YiAPPA,其中T209K/S220E于80 ℃保温20 min后的残留酶活力为37.30%,N237D于80 ℃保温20 min后的残留酶活力为40.98%,T209K/S220E/N237D于80 ℃保温20 min后的残留酶活力为43.15%。以上研究结果显示,通过定点突变引入离子键对植酸酶YiAPPA进行分子改造,获得了绝对酶活不变且热稳定性明显提高的离子键突变体T209K/S220E、N237D及T209K/S220E/N237D。此外,组合突变体T209K/S220E/N237D的热稳定性明显优于突变体T209K/S220E及N237D。已有研究表明,通过组合有益突变进一步改良酶特性的组合突变方法是酶分子定向进化的有效方法[32]。在以上研究结果的基础上,本研究进一步研究了组合突变体T209K/S220E/N237D的酶学特性。
![]() 图 3 重组植酸酶YiAPPA及其突变体的SDS-PAGE分析(A)与酶活和热稳定性检测(B)注:A图中,M:蛋白质marker;1:YiAPPA;2:T209K/S220E;3:N237D;4:T209K/S220E/N237D;B图中不同小写字母代表组间具有显著差异(P<0.05)。Figure 3. SDS-PAGE analysis of recombinant phytase YiAPPA and its mutants (A) and its enzyme activity and thermostability (B)
图 3 重组植酸酶YiAPPA及其突变体的SDS-PAGE分析(A)与酶活和热稳定性检测(B)注:A图中,M:蛋白质marker;1:YiAPPA;2:T209K/S220E;3:N237D;4:T209K/S220E/N237D;B图中不同小写字母代表组间具有显著差异(P<0.05)。Figure 3. SDS-PAGE analysis of recombinant phytase YiAPPA and its mutants (A) and its enzyme activity and thermostability (B)2.2 突变体T209K/S220E/N237D的酶学特性分析
2.2.1 突变体T209K/S220E/N237D的最适反应温度及热力学参数
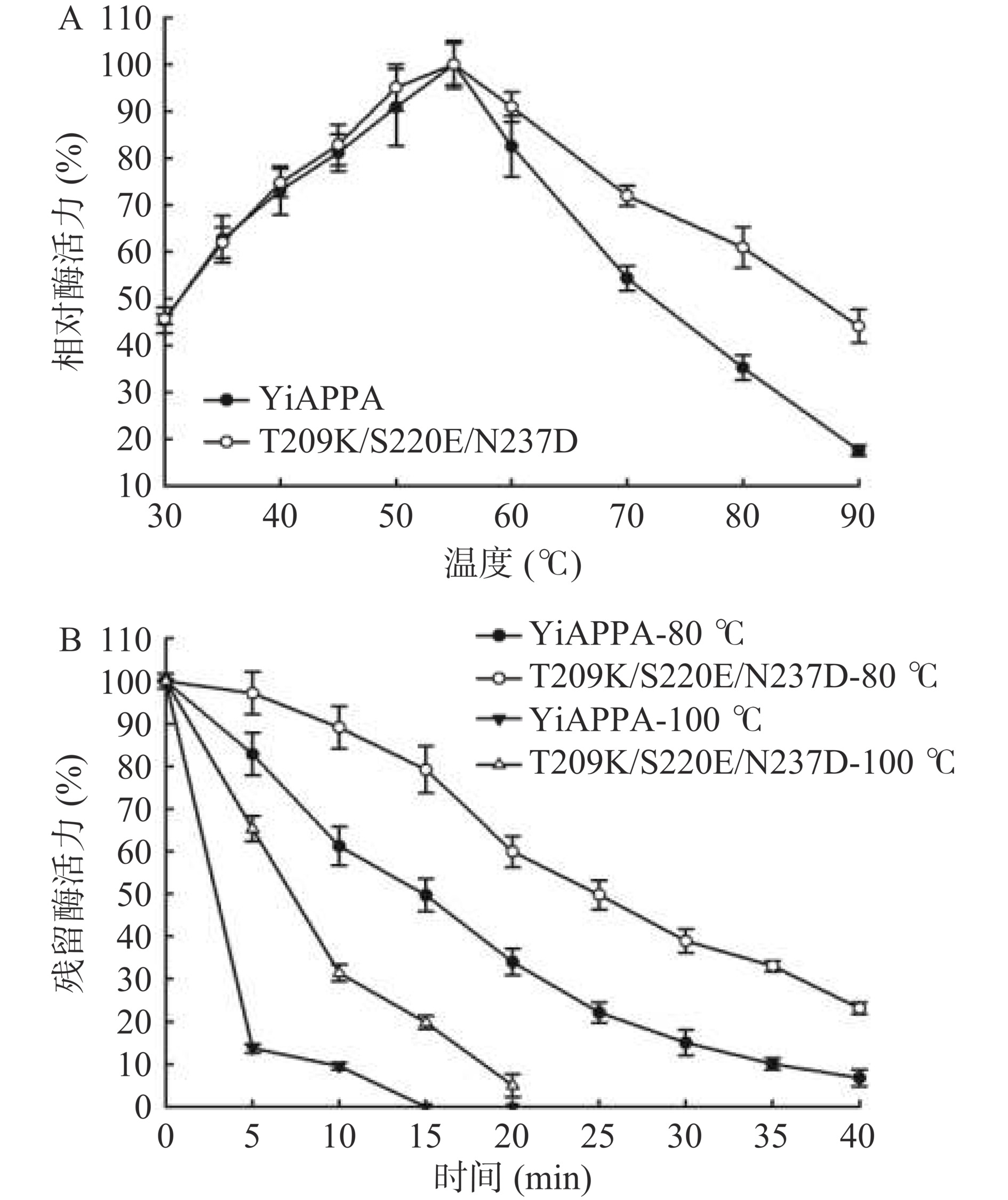
本研究测定了重组植酸酶YiAPPA和突变体T209K/S220E/N237DR的最适反应温度及热力学参数,如图4和表3所示。图4A显示,突变体T209K/S220E/N237D的最适反应温度与YiAPPA一致,均约为55 ℃;在30~55 ℃范围内,T209K/S220E/N237D和YiAPPA相对酶活力基本一致;在60~90 ℃范围内,T209K/S220E/N237D的相对酶活力明显高于YiAPPA的相对酶活力。以上结果表明,突变体T209K/S220E/N237D在较高温度范围内(60~90 ℃)保留有更高的相对酶活力,这可能与其具有更优良的热稳定性相关。图4B显示,突变体T209K/S220E/N237D于80 ℃或100 ℃保温后的残留酶活力显著(P<0.05)高于YiAPPA。T209K/S220E/N237D于80 ℃保温25 min后保留有49.81%的酶活力,YiAPPA于80 ℃保温25 min后仅保留有22.16%的酶活力;T209K/S220E/N237D于100 ℃保温10 min后保留有31.42%的酶活力,YiAPPA于100 ℃保温10 min后仅保留有9.54%的酶活力。
![]() 图 4 温度对重组植酸酶的酶活(A)和稳定性(B)的影响Figure 4. Effects of temperature on enzymatic activity (A) and stability (B) of recombinant phytase表 3 重组植酸酶的热力学参数Table 3. Thermodynamic parameters of recombinant phytase
图 4 温度对重组植酸酶的酶活(A)和稳定性(B)的影响Figure 4. Effects of temperature on enzymatic activity (A) and stability (B) of recombinant phytase表 3 重组植酸酶的热力学参数Table 3. Thermodynamic parameters of recombinant phytase重组植酸酶 80 ℃ t1/2 (min) T5030(℃) Tm(℃) YiAPPA 14.81±0.37 55.12±0.87 48.36±0.53 T209K/S220E/N237D 24.72±0.81 64.05±0.92 55.04±0.59 在以上研究的基础上,进一步测定了重组植酸酶的热力学参数,包括半衰期(t1/2)、半失活温度(T5030)和热解折叠温度(Tm),结果如表3所示。重组植酸酶YiAPPA于80 ℃的半衰期(t1/2)为14.81 min,突变体T209K/S220E/N237D于80 ℃的半衰期(t1/2)为24.72 min,延长了0.67倍;重组YiAPPA的半失活温度(T5030)为55.12 ℃,T209K/S220E/N237D的半失活温度(T5030)为64.05 ℃,提高了8.93 ℃;重组YiAPPA的热解折叠温度(Tm)为48.36 ℃,T209K/S220E/N237D的热解折叠温度(Tm)为55.04 ℃,提高了6.68 ℃。以上结果表明,相较于重组YiAPPA,突变体T209K/S220E/N237D的热稳定性明显提高。
目前已有采用理性设计方法提高植酸酶YiAPPA热稳定性的相关研究。Wang等[33]向植酸酶YiAPPA中引入三个N-糖基化位点,获得了植酸酶活性基本不变而热稳定性大幅提高的突变体M14(A78N/A79Q/G80T/G111N/G113T/G245N/D247S)。突变体M14于80 ℃保温25 min后的残留酶活力达78%,于100 ℃保温10 min后的残留酶活力达75%。Abbasi等[34]将YiAPPA分子表面的氨基酸残基T83突变为R83,获得了热稳定性提高的突变体T83R。T83R于80 ℃保温25 min后的残留酶活力达75%,于100 ℃保温10 min后的残留酶活力达50%,并且其于pH5.15的稳定性提高了71%,但是突变体T83R的植酸酶活性仅为YiAPPA的42.5%。本研究构建得到了植酸酶活性不变并且热稳定性提高的离子键突变体T209K/S220E/N237D。以上研究结果表明,采用不同理性设计方法可以不同程度地提高YiAPPA的热稳定性。并且这些分子改造方法在提高YiAPPA热稳定性的同时,也会影响YiAPPA其他酶学性质(如酶活、pH稳定性等)。因此本研究进一步测定了突变体T209K/S220E/N237D的最适反应pH、pH稳定性、动力学常数及蛋白酶抗性。
2.2.2 突变体T209K/S220E/N237D的最适反应pH及pH稳定性
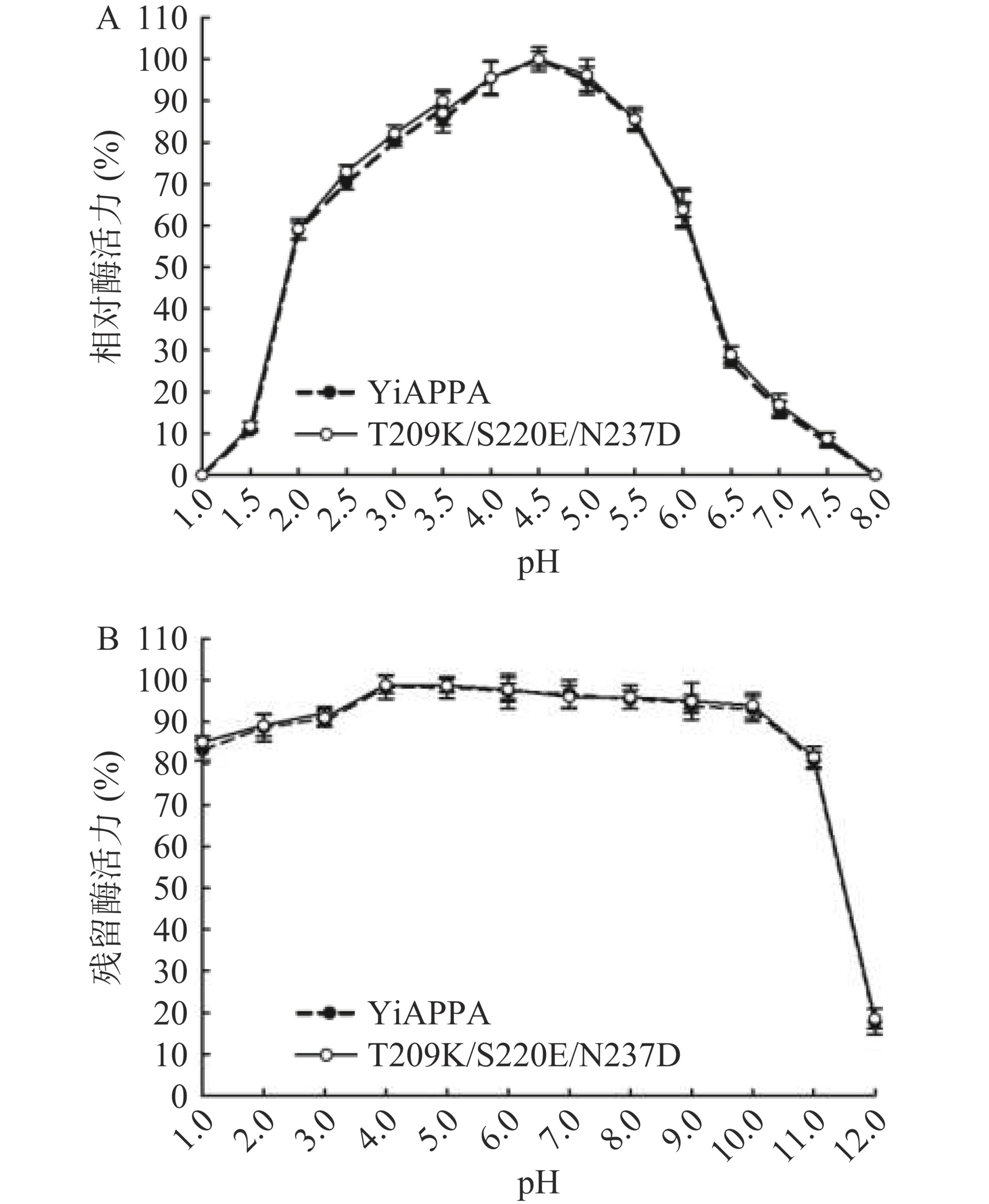
本研究测定了重组植酸酶YiAPPA和突变体T209K/S220E/N237D的最适反应pH及pH稳定性,如图5所示。重组植酸酶YiAPPA和突变体T209K/S220E/N237D的最适反应pH均为4.5,并且两种重组植酸酶在pH1.0~8.0范围内具有相似的相对酶活力(图5A)。此外,重组植酸酶YiAPPA和突变体T209K/S220E/N237D在pH1.0~12.0范围内具有相似的稳定性。如图5B所示,在pH1.0~3.0范围内,两种重组植酸酶均具有80%以上的残留酶活力;在pH4.0~10.0范围内,两种重组植酸酶均具有90%以上的相对酶活;在pH11.0~12.0范围内,两种重组植酸酶均不稳定,相对酶活力迅速下降。以上研究结果表明,向YiAPPA引入离子键K209-E220及D237-R374不影响其最适反应pH及pH稳定性。YiAPPA及突变体T209K/S220E/N237D均具有广泛的pH作用范围,这有利于其在食品工业的应用。
![]() 图 5 pH对重组植酸酶的酶活(A)和稳定性(B)的影响Figure 5. Effects of pH on enzymatic activity (A) and stability (B) of recombinant phytase
图 5 pH对重组植酸酶的酶活(A)和稳定性(B)的影响Figure 5. Effects of pH on enzymatic activity (A) and stability (B) of recombinant phytase2.2.3 突变体T209K/S220E/N237D的动力学参数
重组植酸酶以植酸钠为底物时的绝对酶活及动力学常数如表4所示。突变体T209K/S220E/N237D与重组YiAPPA的绝对酶活基本一致;并且突变体T209K/S220E/N237D的米氏常数Km值和反应常数kcat值也与YiAPPA基本一致。即向YiAPPA引入离子键K209-E220及D237-R374不影响其对底物的结合能力及催化能力。
表 4 重组植酸酶的绝对酶活及动力学常数(37 ℃)Table 4. Specific acitivity and kinetic constants of recombinant phytase (37 ℃)重组植酸酶 绝对酶活(U/mg) Km(μmol/L) kcat(s−1) kcat /Km (μmol/L s−1) YiAPPA 3960.81±105.34 126.24±14.18 10375.27±1256.73 82.19 T209K/S220E/N237D 3982.06±123.09 126.47±12.17 10419.29±1007.16 82.38 通常情况下,酶的活性和稳定性之间存在“代偿效应(trade-off)”[35]。“代偿效应”是改造酶以获得高性能突变酶的限制因素。蛋白质表面改性是对酶活性影响最小的热稳定性提高策略[35]。对酶分子进行蛋白质表面改性有可能克服“代偿效应”,获得稳定性提高、活性不变或者提高的突变酶。酶分子表面离子键对于其维持热稳定性非常重要[36]。通过优化远离活性中心的酶分子表面离子键可以在不影响其催化活性的情况下提高酶稳定性。例如Su等[37]通过定点突变技术向来源于Ochrobactrum sp. M231的甲基对硫磷水解酶MPH_OCH引入分子表面离子键D76-K78,构建得到半失活温度提高4 ℃的突变体P76D/P78K,并且突变体P76D/P78K的催化效率(kcat/Km)提高了2.24倍。Shivange等[38]向来源于Y. mollaretii的植酸酶引入分子表面离子键K77-D122,获得了植酸酶活性基本不变并且热稳定性提高的突变体。该突变体的半失活温度(T5030)提高了1 ℃,其于58 ℃保温20 min后残留酶活力提高了12%。本研究向植酸酶YiAPPA中引入分子表面离子键K209-E220及D237-R374,获得了植酸酶活性、底物结合能力及催化能力基本不变且热稳定性提高的突变体T209K/S220E/N237D。
2.2.4 突变体T209K/S220E/N237D的蛋白酶抗性
重组植酸酶的蛋白酶抗性测定结果如表5所示。重组植酸酶YiAPPA和突变体T209K/S220E/N237D具有相似的蛋白酶抗性。经胃蛋白酶处理后,重组YiAPPA保留有84.17%的残留酶活力,T209K/S220E/N237D保留有85.61%的残留酶活力;经胰蛋白酶处理后,重组YiAPPA保留有82.28%的残留酶活力,T209K/S220E/N237D保留有82.37%的残留酶活力。即向YiAPPA引入离子键K209-E220及D237-R374不影响其蛋白酶抗性。
表 5 重组植酸酶对蛋白酶的抗性(37 ℃,2 h)Table 5. Resistance of recombinant phytase to protease(37 ℃, 2 h)重组植酸酶 胃蛋白酶抗性(%) 胰蛋白酶抗性(%) YiAPPA 84.17±3.20 82.28±4.18 T209K/S220E/N237D 85.61±3.36 82.37±4.02 2.3 重组植酸酶的分子动力学模拟分析
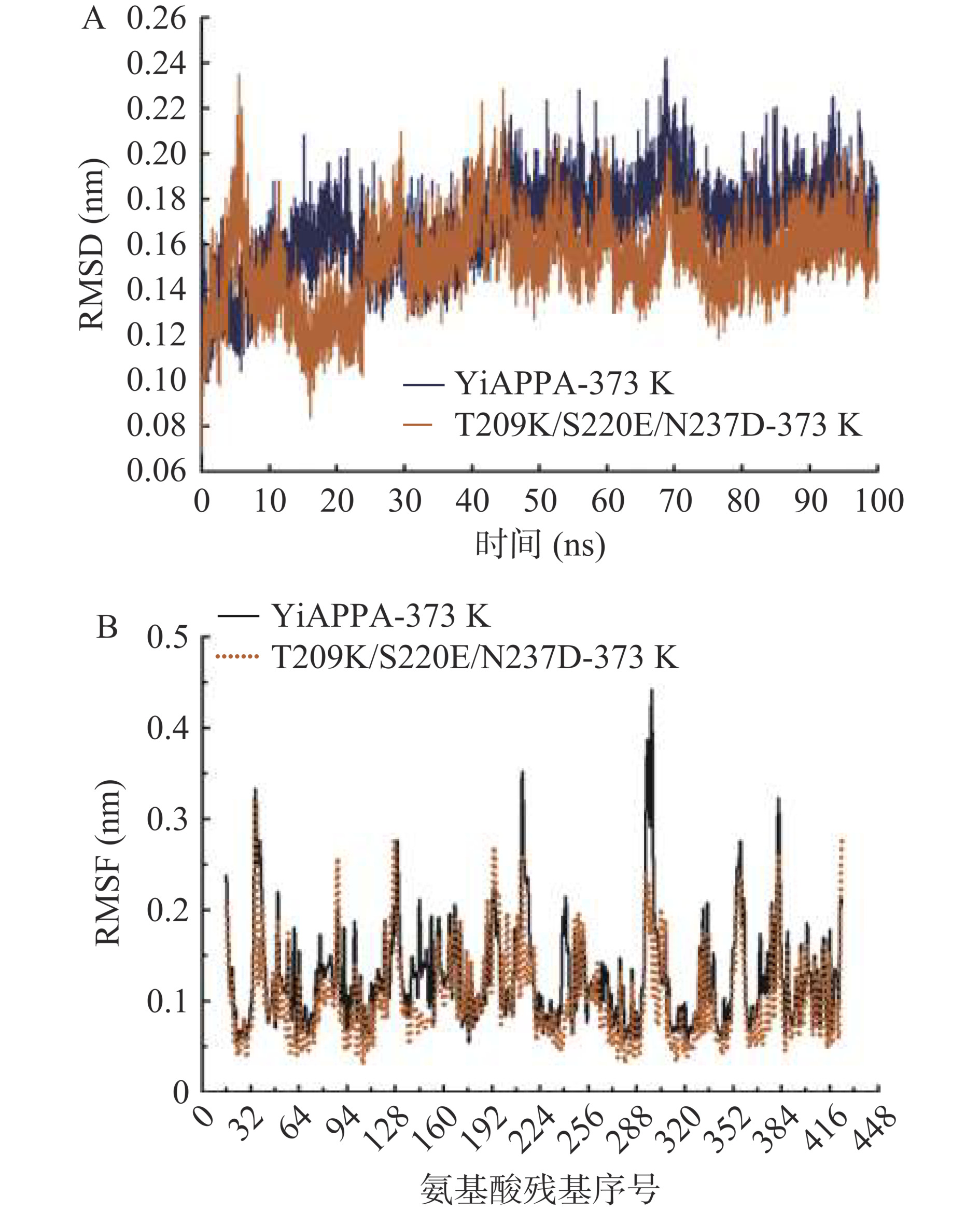
研究表明,蛋白质刚性的增加是蛋白质稳定性增强的主要原因之一,通常表现为较低的RMSD值和RMSF值[39]。为了分析YiAPPA及突变体T209K/S220E/N237D的热力学波动性,本研究对YiAPPA及突变体T209K/S220E/N237D的三级分子结构于373 K下进行100 ns的分子动力学模拟,解析了酶分子整体结构刚性和酶分子中氨基酸残基波动性变化,结果如图6所示。图6A显示,在40 ns后,YiAPPA及突变体T209K/S220E/N237D的RMSD值波动较小,达到平衡,其中YiAPPA的平均RMSD值为0.17 Å,突变体T209K/S220E/N237D的平均RMSD值为0.15 Å。即突变体T209K/S220E/N237D的平均RMSD值低于YiAPPA。酶的RMSD值与其分子整体结构刚性和稳定性相关,酶分子的RMSD值越低,其结构刚性越大,表明其具有更好的稳定性[39]。与YiAPPA相比,突变体T209K/S220E/N237D具有更低的RMSD值,说明突变体T209K/S220E/N237D的分子结构刚性更大,分子结构更稳定。
![]() 图 6 重组植酸酶的分子动力学模拟结果注:A:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSD分析;B:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSF分析。Figure 6. Molecular dynamics simulation results of recombinant phytase
图 6 重组植酸酶的分子动力学模拟结果注:A:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSD分析;B:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSF分析。Figure 6. Molecular dynamics simulation results of recombinant phytase图6B显示,对于突变位点第209位赖氨酸(K209)、突变位点第220位谷氨酸(E220)以及位于这两个突变位点之间的氨基酸残基(第210位缬氨酸至第219位亮氨酸,V210~L219),突变体T209K/S220E/N237D的RMSF值与YiAPPA相比有所降低,这表明YiAPPA中第209位苏氨酸(T209)及第220位丝氨酸(S220)发生突变(T209突变为K209、S220突变为E220)使其以及其周围的氨基酸残基更加稳定,该位点突变同时提高了位点附近氨基酸残基的稳定性。突变体T209K/S220E/N237D中K209、E220以及这两个位点之间的氨基酸残基共同构成β4片层结构-loop结构-β5片层结构(图3),这个结构区域较低的RMSF值表明新引入的离子键K209-E220使得这个结构区域更加稳定。
同样地,对于突变位点第237位天冬氨酸(D237)以及其周围氨基酸残基(第236位谷氨酰胺至第243位天冬氨酸,Q236~D243)、第374位精氨酸(R374)以及其周围氨基酸残基(第375位天冬酰胺至第378位赖氨酸,N375~K378),突变体T209K/S220E/N237D的RMSF值与YiAPPA相比明显降低,这表明YiAPPA中第237位天冬酰胺突变为天冬氨酸(N237D),引入新的离子键D237-R374,同时提高了D237和R374附近区域氨基酸残基的稳定性。突变体T209K/S220E/N237D中D237位于α8-螺旋结构的C端,R374位于α12-螺旋结构的C端,Q236~D243及N375~K378均分别位于紧邻α-螺旋结构的loop结构。以上这些氨基酸残基位点较低的RMSF值表明新引入的离子键D237-R374使得这些结构单元更加稳定。
以上研究结果表明,与YiAPPA相比,突变体T209K/S220E/N237D具有更低的平均RMSD值,并且构成新的离子键的氨基酸残基以及其附近氨基酸残基的RMSF值明显降低。突变体T209K/S220E/N237D的酶分子结构刚性更大,并且其中部分结构单元的稳定性更好,这是突变体T209K/S220E/N237D的热稳定性高于YiAPPA的原因。
3. 结论
本研究通过对YiAPPA中第209位苏氨酸(T209)、第220位丝氨酸(S220)以及第237位天冬酰胺(N237)进行定点突变,引入离子键K209-E220及D237-R374,获得了热稳定性显著提高的突变体T209K/S220E/N237D,其于80 ℃的半衰期(t1/2)为24.72 min,延长了0.67倍;其半失活温度(T5030)为64.05 ℃,提高了8.93 ℃;其热解折叠温度(Tm)为55.04 ℃,提高了6.68 ℃。此外,突变体T209K/S220E/N237D的绝对酶活、动力学参数、最适反应温度、最适反应pH、pH稳定性以及蛋白酶抗性与YiAPPA基本一致。分子动力学模拟结果显示,相较于YiAPPA,突变体T209K/S220E/N237D热稳定性提升的原因在于离子键的引入增强了酶分子结构的刚性,并稳定了其部分结构区域。本研究为植酸酶以及其它类型酶的分子改造提供了重要理论依据,也为YiAPPA在食品加工领域的应用奠定了基础。
-
![]()
图 1 植酸酶YiAPPA(A)和植酸酶rPhyXT52(B)分子表面离子键
注:图中蓝色虚线标记的均为氨基酸残基对之间形成的离子键。
Figure 1. Surficial salt bridges of phytase YiAPPA (A) and phytase rPhyXT52 (B)
![]()
图 2 植酸酶的分子结构分析
注:A:植酸酶YiAPPA的分子结构分析;B:突变体T47E/Q50K/Q64E/T148D/N181D/T209K/S220E/N237D的分子结构分析;图中红色标记的氨基酸残基为突变的氨基酸残基以及与其形成新的离子键的氨基酸残基(Lys209与Glu220形成新的离子键;Asn237与Arg374形成新的离子键);红色虚线标记的为引入的离子键。
Figure 2. Molecular structure analysis of recombinant phytase
![]()
图 3 重组植酸酶YiAPPA及其突变体的SDS-PAGE分析(A)与酶活和热稳定性检测(B)
注:A图中,M:蛋白质marker;1:YiAPPA;2:T209K/S220E;3:N237D;4:T209K/S220E/N237D;B图中不同小写字母代表组间具有显著差异(P<0.05)。
Figure 3. SDS-PAGE analysis of recombinant phytase YiAPPA and its mutants (A) and its enzyme activity and thermostability (B)
![]()
图 4 温度对重组植酸酶的酶活(A)和稳定性(B)的影响
Figure 4. Effects of temperature on enzymatic activity (A) and stability (B) of recombinant phytase
![]()
图 5 pH对重组植酸酶的酶活(A)和稳定性(B)的影响
Figure 5. Effects of pH on enzymatic activity (A) and stability (B) of recombinant phytase
![]()
图 6 重组植酸酶的分子动力学模拟结果
注:A:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSD分析;B:重组植酸酶YiAPPA与T209K/S220E/N237D于373 K(100 ℃)的RMSF分析。
Figure 6. Molecular dynamics simulation results of recombinant phytase
表 1 构建重组质粒所用引物
Table 1 Primers for constructing recombinant plasmids
引物名称 序列 T47E/Q50K-F 5’-GTTACCCCTGACGAGTGGCCGAAATGGC
CGGTCGCCGCAG-3’T47E/Q50K-R 5’-CTGCGGCGACCGGCCATTTCGGCCACTCGT
CAGGGGTAAC-3’Q64E-F 5’-CGAGGTGCAGAATTAGTGACATTGATGG-3’ Q64E-R 5’-CCATCAATGTCACTAATTCTGCACCTCG-3’ T148D-F 5’-GTAAGTTAGATTCGGATCAAACCCATAAGGC-3’ T148D-R 5’-GCCTTATGGGTTTGATCCGAATCTAACTTAC-3’ N181D-F 5’-GATGGGTGAGATTCTGGATTTTGCGGCATCTC-3’ N181D-R 5’-GAGATGCCGCAAAATCCAGAATCTCACCCATC-3’ S220E-F 5’-CAAAAGTCTCGCTCGAAGGACCACTGGCACTG-3’ S220E-R 5’-CAGTGCCAGTGGTCCTTCGAGCGAGACTTTTG-3’ T209K-F 5’-CAGCGAATAAGATCAAGGTGAACAAGCCG-3’ T209K-R 5’-CGGCTTGTTCACCTTGATCTTATTCGCTG-3’ N237D-F 5’-GATCTTTTTGCTACAAGATTCACAAGCGATGC-3’ N237D-R 5’-GCATCGCTTGTGAATCTTGTAGCAAAAAGATC-3’ 注:方框标注的部分为突变的碱基。  下载: 导出CSV
下载: 导出CSV
表 2 植酸酶rPhyXT52分子表面离子键与植酸酶YiAPPA分子表面离子键对比分析
Table 2 Comparative analysis of surficial salt bridges between phytase rPhyXT52 and phytase YiAPPA
rPhyXT52分子
表面
离子键rPhyXT52分子表面离子键及相关
氨基酸残基
在YiAPPA中对应情况YiAPPA分子
表面
离子键Glu45-Lys48 无对应离子键,Thr47-Gln50 Asp46-Arg248 Glu62-Arg59 无对应离子键,Gln64-Arg61 Asp127-Lys130 Asp126-Lys129 有对应离子键,Asp127-Lys130 Glu156-Lys152 Asp147-Lys151 无对应离子键,Thr148-Lys152 Glu164-Lys167 Asp180-Arg247 无对应离子键,Gln181-Arg248 Glu231-Arg31 Glu219-Lys208 无对应离子键,Ser220-Thr209 Glu288-Arg295 Glu230-Arg29 有对应离子键,Glu231-Arg31 Asp338-Arg27 Asp236-Arg372 无对应离子键,Asn237-Arg374 Glu419-Arg277 Asp336-Arg25 有对应离子键,Asp338-Arg27 注:下划线标注的是YiAPPA中拟进行定点突变的氨基酸残基。
下载: 导出CSV
表 3 重组植酸酶的热力学参数
Table 3 Thermodynamic parameters of recombinant phytase
重组植酸酶 80 ℃ t1/2 (min) T5030(℃) Tm(℃) YiAPPA 14.81±0.37 55.12±0.87 48.36±0.53 T209K/S220E/N237D 24.72±0.81 64.05±0.92 55.04±0.59
下载: 导出CSV
表 4 重组植酸酶的绝对酶活及动力学常数(37 ℃)
Table 4 Specific acitivity and kinetic constants of recombinant phytase (37 ℃)
重组植酸酶 绝对酶活(U/mg) Km(μmol/L) kcat(s−1) kcat /Km (μmol/L s−1) YiAPPA 3960.81±105.34 126.24±14.18 10375.27±1256.73 82.19 T209K/S220E/N237D 3982.06±123.09 126.47±12.17 10419.29±1007.16 82.38
下载: 导出CSV
表 5 重组植酸酶对蛋白酶的抗性(37 ℃,2 h)
Table 5 Resistance of recombinant phytase to protease(37 ℃, 2 h)
重组植酸酶 胃蛋白酶抗性(%) 胰蛋白酶抗性(%) YiAPPA 84.17±3.20 82.28±4.18 T209K/S220E/N237D 85.61±3.36 82.37±4.02
下载: 导出CSV
-
[1] PHILIPPI H, SOMMERFELD V, WINDISCH W, et al. Interactions of zinc with phytate and phytase in the digestive tract of poultry and pigs:a review[J]. Journal of the Science of Food and Agriculture,2023,103(15):7333−7342. doi: 10.1002/jsfa.12879
[2] HANDA V, SHARMA D, KAUR A, et al. Biotechnological applications of microbial phytase and phytic acid in food and feed industries[J]. Biocatalysis and Agricultural Biotechnology,2020,25:101600. doi: 10.1016/j.bcab.2020.101600
[3] HUSSAIN S M, HANIF S, SHARIF A, et al. Unrevealing the sources and catalytic functions of phytase with multipurpose characteristics[J]. Catalysis Letters,2021,152(5):1−14.
[4] RIZWANUDDIN S, KUMAR V, NAIK B, et al. Microbial phytase:their sources, production, and role in the enhancement of nutritional aspects of food and feed additives[J]. Journal of Agriculture and Food Research, 2023:100559.
[5] SONG H Y, EL SHEIKHA A F, HU D M. The positive impacts of microbial phytase on its nutritional applications[J]. Trends in Food Science & Technology,2019,86:553−562.
[6] SINGH B, KUMAR G, KUMAR V, et al. Enhanced phytase production by Bacillus subtilis subsp. subtilis in solid state fermentation and its utility in improving food nutrition[J]. Protein and Peptide Letters,2021,28(10):1083−1089. doi: 10.2174/0929866528666210720142359
[7] YAVER E. Dephytinized flaxseed flours by phytase enzyme and fermentation:Functional ingredients to enhance the nutritional quality of noodles[J]. Journal of the Science of Food and Agriculture,2023,103(4):1946−1953. doi: 10.1002/jsfa.12266
[8] PRAGYA, SHARMA K K, KUMAR S, et al. Enhanced production and immobilization of phytase from Aspergillus oryzae:A safe and ideal food supplement for improving nutrition[J]. Letters in Applied Microbiology, 2023, 76(2).
[9] THAKUR N, PATEL S K S, KUMAR P, et al. Bioprocess for hyperactive thermotolerant Aspergillus fumigatus phytase and its application in dephytinization of wheat flour[J]. Catalysis Letters,2022,152(11):3220−3232. doi: 10.1007/s10562-021-03886-0
[10] TRUELOCK C N, YODER A D, EVANS C E, et al. The effects of pelleting process parameters and phytase source on the in-feed stability of phytase[J]. Animal Feed Science and Technology,2022,294:115407. doi: 10.1016/j.anifeedsci.2022.115407
[11] SHIVANGE A V, SCHWANEBERG U. Recent advances in directed phytase evolution and rational phytase engineering[J]. Directed Enzyme Evolution:Advances and Applications, 2017:145−172.
[12] VASUDEVAN U M, JAISWAL A K, KRISHNA S, et al. Thermostable phytase in feed and fuel industries[J]. Bioresource Technology,2019,278:400−407. doi: 10.1016/j.biortech.2019.01.065
[13] USHASREE M V, SHYAM K, VIDYA J, et al. Microbial phytase:impact of advances in genetic engineering in revolutionizing its properties and applications[J]. Bioresource Technology,2017,245:1790−1799. doi: 10.1016/j.biortech.2017.05.060
[14] LIAO Y, LI C M, CHEN H, et al. Site-directed mutagenesis improves the thermostability and catalytic efficiency of Aspergillus niger N25 phytase mutated by I44E and T252R[J]. Applied Biochemistry and Biotechnology,2013,171(4):900−915. doi: 10.1007/s12010-013-0380-2
[15] ZHANG Z J, YANG J, XIE P J, et al. Characterization of a thermostable phytase from Bacillus licheniformis WHU and further stabilization of the enzyme through disulfide bond engineering[J]. Enzyme and Microbial Technology,2020,142:109679. doi: 10.1016/j.enzmictec.2020.109679
[16] WANG X, DU J, ZHANG Z Y, et al. A rational design to enhance the resistance of Escherichia coli phytase appA to trypsin[J]. Applied Microbiology and Biotechnology,2018,102:9647−9656. doi: 10.1007/s00253-018-9327-4
[17] LI J D, LI X L, GAI Y M, et al. Evolution of E. coli phytase for increased thermostability guided by rational parameters[J]. Journal of Microbiology and Biotechnology,2019,29(3):419−428. doi: 10.4014/jmb.1811.11017
[18] HUANG H Q, LUO H Y, YANG P L, et al. A novel phytase with preferable characteristics from Yersinia intermedia[J]. Biochemical and Biophysical Research Communications,2006,350(4):884−889. doi: 10.1016/j.bbrc.2006.09.118
[19] 袁林, 黄朝, 曾静, 等. 植酸酶YiAPPA与生淀粉结合域SBD融合酶的构建及酶学性质分析[J]. 生物技术通报,2018,34(3):200−207. [YUAN L, HUANG Z, ZENG J, et al. Fusion of Phytase YiAPPA with the Raw-starch Binding Domain and Characterization of the Fusion Enzyme[J]. Biotechnology Bulletin,2018,34(3):200−207.] YUAN L, HUANG Z, ZENG J, et al. Fusion of Phytase YiAPPA with the Raw-starch Binding Domain and Characterization of the Fusion Enzyme[J]. Biotechnology Bulletin, 2018, 34(3): 200−207.
[20] TAN H, WU X, XIE L Y, et al. Identification and characterization of a mesophilic phytase highly resilient to high-temperatures from a fungus-garden associated metagenome[J]. Applied Microbiology and Biotechnology,2016,100(5):2225−2241. doi: 10.1007/s00253-015-7097-9
[21] NEZHAD N G, RAJA A R R N Z, NORMI Y M, et al. Integrative structural and computational biology of phytases for the animal feed industry[J]. Catalysts,2020,10(8):844. doi: 10.3390/catal10080844
[22] FU D W, HUANG H Q, LUO H Y, et al. A highly pH-stable phytase from Yersinia kristeensenii:cloning, expression, and characterization[J]. Enzyme and Microbial Technology,2008,42(6):499−505. doi: 10.1016/j.enzmictec.2008.01.014
[23] WATERHOUSE A, BERTONI M, BIENERT S, et al. SWISS-MODEL:Homology modelling of protein structures and complexes[J]. Nucleic Acids Research,2018,46(W1):W296−W303. doi: 10.1093/nar/gky427
[24] PIOVESAN D, MINERVINI G, TOSATTO S C E. The RING 2.0 web server for high quality residue interaction networks[J]. Nucleic Acids Research,2016,44(W1):W367−W374. doi: 10.1093/nar/gkw315
[25] ANAGNOSTOPOULOS C, SPIZIZEN J. Requirements for transformation in Bacillus subtilis[J]. Journal of Bacteriology,1961,81(5):741. doi: 10.1128/jb.81.5.741-746.1961
[26] GREEN M R, SAMBROOK J. Molecular cloning:a laboratory manual[M]. New York:Cold Spring Harbor Laboratory Press, 2012:101−200.
[27] LAEMMLI U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J]. Nature,1970,227:680−685. doi: 10.1038/227680a0
[28] BUSTO M D, APENTEN R K O, ROBINSON D S, et al. Kinetics of thermal inactivation of pea seed lipoxygenases and the effect of additives on their thermostability[J]. Food Chemistry,1999,65(3):323−329. doi: 10.1016/S0308-8146(98)00216-7
[29] LIM J H, HWANG K Y, CHOI J, et al. Mutational effects on thermostable superoxide dismutase from Aquifex pyrophilus:understanding the molecular basis of protein thermostability[J]. Biochemical and Biophysical Research Communications,2001,288(1):263−268. doi: 10.1006/bbrc.2001.5752
[30] FJELLSTEDT T A, SCHLESSELMAN J J. A simple statistical method for use in kinetic analysis based on Lineweaver-Burk plots[J]. Analytical Biochemistry,1977,80(1):224−238. doi: 10.1016/0003-2697(77)90641-8
[31] VAN D S D, LINDAHL E, HESS B, et al. GROMACS:fast, flexible, and free[J]. Journal of Computational Chemistry, 2005, 26(16):1701−1718.
[32] 蔡婷婷, 曹佳仁, 邱帅, 等. 半理性设计进化土曲霉来源的ω-转氨酶AtTA热稳定性[J]. 生物工程学报,2023,39(6):2126−2140. [CAI T T, CAO J R, QIU S, et al. Semi-rational evolution of ω-transaminase from Aspergillus terreus for enhancing the thermostability[J]. Chinese Journal of Biotechnology,2023,39(6):2126−2140.] CAI T T, CAO J R, QIU S, et al. Semi-rational evolution of ω-transaminase from Aspergillus terreus for enhancing the thermostability[J]. Chinese Journal of Biotechnology, 2023, 39(6): 2126−2140.
[33] WANG Q, LIU X Q, TIAN J, et al. Enhancing the thermostability of phytase to boiling point by evolution-guided design[J]. Applied and Environmental Microbiology,2022,88(11):e00506.
[34] ABBASI K M, SAFFAR B, HEMMATI R, et al. Thermally stable and acidic pH tolerant mutant phytases with high catalytic efficiency from Yersinia intermedia for potential application in feed industries[J]. Environmental Science and Pollution Research,2022,29(22):33713−33724. doi: 10.1007/s11356-022-18578-4
[35] SIDDIQUI K S. Defying the activity–stability trade-off in enzymes:taking advantage of entropy to enhance activity and thermostability[J]. Critical Reviews in Biotechnology,2017,37(3):309−322. doi: 10.3109/07388551.2016.1144045
[36] XU Z, CEN Y K, ZOU S P, et al. Recent advances in the improvement of enzyme thermostability by structure modification[J]. Critical Reviews in Biotechnology,2020,40(1):83−98. doi: 10.1080/07388551.2019.1682963
[37] SU Y D, TIAN J, WANG P, et al. Improving the thermostability of a methyl parathion hydrolase by adding the ionic bond on protein surface[J]. Applied Biochemistry and Biotechnology,2011,165:989−997. doi: 10.1007/s12010-011-9314-z
[38] SHIVANGE A V, ROCCATANO D, SCHWANEBERG U. Iterative key-residues interrogation of a phytase with thermostability increasing substitutions identified in directed evolution[J]. Applied Microbiology and Biotechnology,2016,100(1):227−242. doi: 10.1007/s00253-015-6959-5
[39] BENSON N C, DAGGETT V. A comparison of multiscale methods for the analysis of molecular dynamics simulations[J]. The Journal of Physical Chemistry B,2012,116(29):8722−8731. doi: 10.1021/jp302103t
 下载:
下载:






计量
- 文章访问数: 65
- HTML全文浏览量: 6
- PDF下载量: 11
