


Gene Expression, Bioinformatics and Substrate Selectivity of Phospholipase D from Acinetobacter sp.
-
摘要: 微生物来源磷脂酶D(Phospholipase D,PLD)因其较高的催化活性和广泛的底物选择谱而成为磷脂合成应用中的热点。本研究以Acinetobacter sp. DUT-2来源的PLD(ADPLD)为研究对象,首先通过生物信息学分析蛋白序列特征,然后构建重组质粒并在大肠杆菌中实现异源表达,进一步纯化酶蛋白并分析ADPLD对不同酰基链长磷脂酰胆碱(PC)的底物选择性,最后通过分子对接和分子模拟探究ADPLD的底物识别机制。通过对ADPLD和其他微生物来源的PLD多序列比对和进化树分析表明,ADPLD与链霉菌来源PLD序列相似度低于30%,且只有一个保守的HKD基序,这表明ADPLD的催化机制可能与传统认知中需要两个HKD基序完成PLD催化过程的反应机制有所不同。ADPLD主要以可溶性蛋白形式表达,仅通过Ni2+亲和层析在50 mmol/L的低浓度咪唑下便能纯化出较为均一的蛋白,以大豆PC为底物时的比活性约为4.09 U/mg。ADPLD对中和短链PC(C6~C14)的活性相对较高,其中ADPLD对8:0/8:0-PC的比活性高于其它酰基链长的PC底物,为13.2 U/mg。当PC的酰基链长从C14增加至C16时,ADPLD对PC的活性明显降低。分子模拟和分子对接结果显示ADPLD的氨基酸残基Thr205、Pro209、Phe293、Ala324、Lys329和Phe453能够分别与PC形成疏水相互作用。Arg383和Gly326能够与PC形成氢键,其中Arg383(N)、Gly326(N)与PC(P)之间的距离<3 Å。这些结果表明,ADPLD能够与磷脂分子形成一个稳定的酶与底物的中间体。研究结果为ADPLD的分子改造和进一步工业应用奠定了基础。Abstract: Microbial phospholipase D (PLD) showed a more competent potential for phospholipids production due to its higher catalytic activity and broader substrate specificity. In this study, the PLD from Acinetobacter sp. DUT-2 (ADPLD) was used as the research object. Firstly, bioinformatics was used to examine the protein sequence characteristics. Then, the recombinant plasmid was generated and heterologously expressed in Escherichia coli. The enzyme protein was purified further, and the substrate selectivity of ADPLD to phosphatidylcholine (PC) with various acyl chains was investigated. Finally, the substrate recognition mechanism of ADPLD was investigated using molecular docking and molecular dynamics simulation. Multiple sequence alignment and phylogenetic tree analysis of ADPLD and other microbial-derived PLDs revealed that the sequence similarity between ADPLD and Streptomyces-derived PLDs was less than 30%, and there was only one conserved HKD motif, indicating that the catalytic mechanism of ADPLD might differ from the reaction mechanism in traditional cognition, which required two HKD motifs to complete the PLD catalytic process. ADPLD was mostly produced as a soluble protein, and a relatively uniform protein could be purified using Ni2+ affinity chromatography at a low concentration of 50 mmol/L imidazole. When soybean PC was utilized as a substrate, the specific activity of ADPLD was about 4.09 U/mg. ADPLD showed relatively high activity in neutralizing short-chain PC (C6~C14), with a specific activity of 13.2 U/mg for 8:0/8:0-PC, which was higher than that of other PC substrates with long acyl chains. The activity of ADPLD on PC reduced dramatically when the acyl chain length of PC grew from C14 to C16. Molecular dynamics simulation and molecular docking experiments revealed that the ADPLD amino acid residues Thr205, Pro209, Phe293, Ala324, Lys329, and Phe453 might form hydrophobic interactions with PC. Arg383 and Gly326 could form hydrogen bonds with PC when the distance between Arg383 (N), Gly326 (N), and PC (P) was <3 Å. These results indicate that ADPLD can form a stable enzyme substrate intermediate with phospholipid molecules. These findings set the groundwork for ADPLD molecular modification and future industrial applications.
-
Keywords:
- Acinetobacter /
- phospholipase D /
- enzymatic property /
- protein expression /
- phospholipid
-
磷脂是所有生物体中生物膜的主要组成成分[1],其主要消费市场是食品和饮料领域。我国是磷脂的主要消费地区,2018年我国的消费量占全球消费总量的27.70%。2018年,全球卵磷脂的产量为58.3万吨,在发展中国家快速增长的需求下,预计到2025年将增长到85.5万吨。因此,具有多种生理功能的磷脂在食品、化妆品以及制药工业中具有很大的市场潜力[2−3]。
磷脂酶D(PLD,EC 3.1.4.4)是一类水解磷脂生成磷脂酸和羟基化合物的酶。此外,PLD还可催化磷脂极性头部基团的转移反应,从而将自然界中含量丰富的磷脂酰胆碱(PC)催化合成其他稀有磷脂[4]。PLD酶转化法的反应条件温和,高效易控。伴随着近几年绿色生物质溶剂的快速发展,以酶制剂为核心的绿色、低碳、高效生物加工成为国际农产品加工领域的发展趋势[5]。目前报道指出微生物来源PLD表现出更加广泛的底物特异性和催化活性[6]。近年来,人们开发了多种方法用来挖掘具有潜在工业应用价值的微生物PLD,其中传统筛选和宏基因组策略得到了广泛的应用。然而,这些方法需要筛选大量的菌株,并且方法的靶标不明确,往往难以获得高催化活性的PLD酶蛋白。随着基因数量呈指数级增长,基因组测序和注释在数据库中得到应用,基因组挖掘近年来备受关注。研究人员可以从 GenBank、Pfam、Brenda 等各种公共数据库中检索出已定义功能的,具有所需属性的基因。Zhou等[7]基于基因组测序和注释,建立了氨基酸序列相似度、保守基序和生物来源安全性的3个筛选标准。基于这3个筛选条件,成功挖掘了一种来源于Streptomyces mobaraensis的PLD,该PLD可以催化大豆卵磷脂和L-丝氨酸合成磷脂酰丝氨酸(PS)。截至目前,链霉菌[8]、芽孢杆菌[9]、大肠杆菌[10]、不动杆菌[11]、苍白杆菌[12]、铜绿假单胞菌[13]、鼠伤寒沙门氏菌[14]和棒状杆菌[15]等微生物来源的PLD已被报道[16],其中Mao等[11]报道不动杆菌(Acinetobacter radioresistens a2)PLD催化的转磷脂酰化反应对合成磷脂酰丝氨酸(PS)具有较高转化率和选择性。这些研究提示微生物来源的PLD(特别是不动杆菌PLD)可能具有更好的稀有磷脂合成潜力,但与PLD超家族大量的序列信息相比,酶学性质信息的极度缺乏严重阻碍了对PLD酶结构-功能关系的深入了解及该酶的进一步应用。若能扩大PLD的来源,挖掘具有相对更高催化活性和纯化步骤简单的微生物来源PLD,可助力解决功能食品开发对高品质磷脂的迫切需求,推动我国食品产业的发展。
本研究从NCBI数据库中挖掘了一个来源于Acinetobacter sp. DUT-2 的PLD(ADPLD)。依据大肠杆菌的密码子偏好性设计编码ADPLD的基因序列,构建ADPLD的大肠杆菌表达载体,并导入大肠杆菌SHuffle T7中进行异源表达;利用Ni亲和层析分离纯化ADPLD蛋白,采用分子模拟和分子对接方法研究ADPLD底物识别的分子基础。
1. 材料与方法
1.1 材料与仪器
大肠杆菌SHuffle T7感受态细胞 购自上海唯地生物技术有限公司;pET-28a载体质粒 购自Invitrogen公司;ADPLD成熟肽基因、PCR引物 由生工生物工程(上海)有限公司合成;T4 DNA连接酶(350 U/μL)、限制性内切酶(EcoR I,Not I)(1 μg λ DNA/μL)、高保真PCR聚合酶(5 sec/kb)、琼脂糖、dNTPs 均购自TaKaRA生物技术(北京)有限公司;辣根过氧化物酶(300 U/mg)、蛋白中分子量范围Marker、质粒DNA抽提试剂盒、PCR产物纯化试剂盒、BCA蛋白质定量检测试剂盒 均购自生工生物工程(上海)有限公司;卡那霉素(Kanamycin)、三羟甲基甲烷(Tris)、异丙基硫代半乳糖苷(IPTG)、甘氨酸(Gly)、酵母提取物、蛋白胨 均购自Sigma-Aldrich公司;大豆PC(纯度95%) 购自Avanti Polar Lipids公司;胆碱酶(4 U/mg) 由华南理工大学工业酶与生物脂质实验室制备与保存[17]。
CB 800 V超净工作台 苏洁医疗器械苏州有限公司;K-300蛋白电泳仪 北京科创锐新生物科技有限公司;ETC811 PCR仪 苏州东胜兴业科学仪器有限公司;JS-780全自动凝胶成像仪 上海培清科技有限公司;QuickSpeed 4000小型高速离心机 莫纳生物科技有限公司;LDZF-50KB灭菌锅 上海申安医疗器械厂;ZS-N生化培养箱 浙江华盛源仪器有限公司;BSA224S分析天平 赛多利斯科学仪器有限公司;PHS-3C pH计 上海仪电科学仪器股份有限公司;JY96-IIN超声破碎仪 上海沪析仪器有限公司;H4-20KR高速冷冻离心机 湖南可成仪器设备有限公司;SPARK多功能酶标仪Infinite MNANO TECAN公司。
1.2 实验方法
1.2.1 ADPLD基因序列的生物信息学分析
从NCBI数据库中下载不动杆菌Acinetobacter sp. DUT-2 PLD(ADPLD)的完整氨基酸序列(GenBank:AMO39720.1)。采用Signal-5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)分析其信号肽序列。采用ExPASy-PROSITE(https://prosite.expasy.org/)分析其活性位点。采用ProtParam tool(https://web.expasy.org/protparam/)分析ADPLD成熟肽氨基酸组成、分子量及等电点等理化性质[18]。采用Multalin在线分析系统(http://multalin.toulouse.inra.fr/multalin/)和NCBI中的BLAST工具将ADPLD的氨基酸序列与NCBI数据库中记录的且通过实验证明具有明确PLD活性的蛋白同源序列进行比对。基于泊松修正模型(Poisson correction)的邻接法(Neighbor joining method),采用MEGA X软件构建系统发育进化树。用自举检验(Bootstrap test)估算系统进化树各分支节点的置信度,累积共1000次循环[19−21]。
1.2.2 ADPLD基因的克隆
根据大肠杆菌密码子偏好性优化ADPLD的成熟肽基因序列。分别将限制性内切酶EcoRI和NotI酶切位点添加在优化后的基因序列上下游。ADPLD成熟肽的基因序列由生工生物工程(上海)有限公司进行全基因合成,并插入到pET-28a载体中。用软件Primer Premier 5.0设计特异性引物进行PCR扩增,引物ADPLD-F的序列为:GAATTCATCTACCACACCGTGAAACC;引物ADPLD-R的序列为:GCGGCCGCGAAGGTGCTCAG(下划线部分分别为EcoRI和NotI酶切位点)。PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s;55 ℃退火30 s;72 ℃延伸1 min 30 s;30个循环;最终72 ℃延伸10 min。将PCR产物进行1%琼脂糖凝胶电泳鉴定,使用PCR产物纯化试剂盒回收目的基因。
1.2.3 ADPLD酶蛋白的发酵表达
将pET-28a-ADPLD重组质粒通过热激法转化至大肠杆菌SHuffle T7感受态细胞中,将菌液涂布于含有50 μg/mL卡那霉素的肉膏蛋白胨(LB)固体平板上,在37 ℃恒温培养箱中过夜培养。将菌落PCR验证过的阳性克隆子接种于含有50 μg/mL卡那霉素的5 mL LB液体培养基中,在37 ℃、200 r/min条件下过夜培养,作为一级种子液。向含有500 mL LB液体培养基的2 L锥形瓶中接入1%的一级种子液扩大培养,在37 ℃,200 r/min条件下培养至菌体OD600为0.6,将摇床培养温度冷却至16 ℃,于超净台中将500 μL的1 mol/L异丙基-β-D-硫代半乳糖苷(IPTG)加入到该2 L锥形瓶中(终浓度为1 mmol/L),然后在16 ℃,200 r/min条件下继续诱导12 h。通过冷冻离心机在4 ℃,4000 r/min条件下离心10 min分离发酵液和菌体,收集菌体备用。
1.2.4 ADPLD酶蛋白的纯化
称取适量菌体加入到菌体湿重10倍质量的Ni柱(Ni-NTA琼脂糖纯化树脂)纯化缓冲液A(20 mmol/L Tris-HCl,20 mmol/L咪唑,100 mmol/L NaCl,pH8.0)中,充分搅拌混匀。在功率70%,破碎2 s,停止2 s条件下超声破碎20 min。超声破碎后12000 r/min条件下离心20 min获得ADPLD粗酶液。用Ni柱纯化缓冲液A平衡Ni柱,然后将经0.45 μm滤膜过滤的粗酶液以3 mL/min流速缓慢加样到Ni柱中,多次穿过,收集穿过液。待上样结束后,用5倍柱体积的Ni柱纯化缓冲液A冲洗平衡Ni柱,接着用不同咪唑浓度的Ni柱缓冲液以2倍体积,5 mL/min流速洗脱Ni柱,收集各咪唑浓度的洗脱液。测定各组分蛋白浓度,十二烷基硫酸钠聚丙烯酰胺凝胶(SDS-PAGE)蛋白电泳检测目标蛋白。
1.2.5 PLD水解活性测定
采用酶联比色法测定PLD的水解活性[22]。反应体系为100 μL Tris-HCl(100 mmol/L,pH8.0)缓冲液中包含0.4 mmol/L大豆PC或不同酰基链长的PC底物(C6~C18),20 mmol/L CaCl2和10 μL纯酶样品。在40 ℃振荡孵育10 min后,沸水加热反应体系使酶蛋白变性失活,冷却至室温后,加入包含50 mmol/L 乙二胺四乙酸(EDTA),42 mmol/L苯酚,50 mmol/L 4-氨基安替比林,0.3 U过氧化酶和0.111 U胆碱氧化酶的50 μL Tris-HCl缓冲液(1 mol/L,pH8.0)终止反应并显色。在37 ℃孵育60 min,500 nm处测定反应混合物的吸光度。对照实验采用10 μL蒸馏水代替酶溶液,在同样条件下处理反应体系。用不同浓度的氯化胆碱标准溶液代替反应体系制作标准曲线。PLD水解活性的定义为:以大豆PC为底物,每分钟催化底物水解产生1 μmol胆碱所需要的酶量为1个单位(U)的PLD水解活力。
1.2.6 分子模拟
使用I-TASSER(https://zhanggroup.org//I-TASSER/)在线系统对ADPLD进行建模[23−25]。使用PROCHECK[26]和QMEAN[27]在线系统对预测的ADPLD三维模型进行验证和质量评估。利用AutoDock tools软件建立小分子配体和ADPLD蛋白的pdbqt文件[28]。利用AutoDock tools软件构建ADPLD与底物的复合结构。选择评分最高的ADPLD与底物的复合结构作为构象,进行后续分析。使用Packmol软件创建PC单层膜模型[29]。采用GROMACS(Groningen machine for molecular dynamics simulations)和Charmm36力场进行膜蛋白分子动力学模拟[30]。使用Protein-Ligand Interaction Profiler(https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index)在线系统、PyMOL和LigPlus软件对ADPLD与底物的复合结构间的相互作用可视化[31−34]。
1.3 数据处理
采用SPSS软件进行ANOVA统计分析,结果以平均值±标准差表示,采用Origin软件绘图。
2. 结果与分析
2.1 ADPLD编码氨基酸序列的比对分析
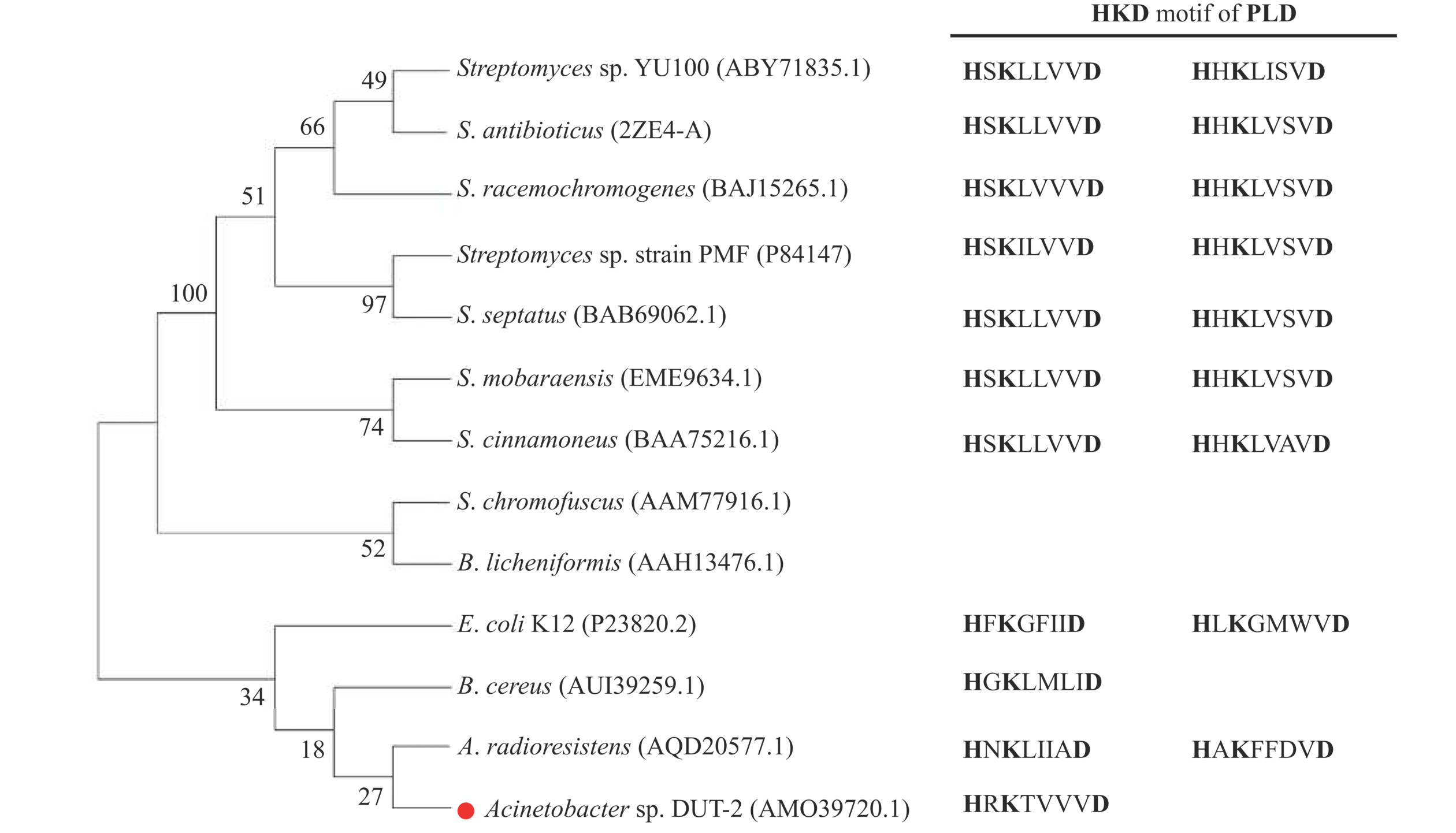
比较了目前已经被实验证实具有活性的PLD酶蛋白的氨基酸序列。如图1所示,根据系统进化树分析,可以将鉴定结果分为两类,第一类PLD主要来源于链霉菌,显示出较高的氨基酸序列相似性,主要包括Streptomyces sp. YU100(GenBank accession No. ABY71835.1),S. antibioticus (2ZE4-A),S. racemochromogenes(BAJ15265.1),Streptomyces sp. strain PMF(P84147),S. septatus(BAB69062.1),S. mobaraensis(EME96334.1)和S. cinnamoneus(BAA75216.1)。截至目前,除S. chromofuscus外,其他链霉菌来源的PLD都具有两个严格保守的HKD基序[35]。另一类PLD主要来源于S. chromofuscus(AAM77916.1),B. licheniformis(AAH13476.1),E. coli K12(P23830.2),B. cereus(AUI39259.1),A. radioresistens(AQD20577.1)和Acinetobacter sp. DUT-2(AMO39720.1),该类PLD与链霉菌来源PLD的序列相似度低(<70%)。此外,后者通过进一步序列分析发现B. licheniformis和S. chromofuscus来源的PLD均没有任何保守的HKD基序[36],而B. cereus来源的PLD有一个HKD基序。E. coli K12和A. radioresistens来源的PLD虽然都具有两个HKD基序,但与链霉菌PLD的序列相似性低,氨基酸组成差异大。值得注意的是,本研究中报道的Acinetobacter sp. DUT-2 PLD与A. radioresistens(AQD20577.1)的PLD虽均来源于不动杆菌,但Acinetobacter sp. DUT-2 来源的PLD只有一个HKD基序。目前主流观点认为,PLD超家族的两个HKD基序构成了PLD的活性位点。在反应过程中,一个HKD基序上的组氨酸亲核进攻底物磷脂的P-O键,形成磷脂酰组氨酸中间体,然后另一个HKD基序上的组氨酸提供一个氢原子,对进入的醇去质子化,活化的醇作为第二亲核体攻击磷脂酰组氨酸中间体从而产生新的磷脂产物。单一的HKD基序可能预示着ADPLD在底物识别和催化活性上具有区别于拥有两个HKD基序PLD的酶学特征[37]。
![]() 图 1 微生物PLD氨基酸序列的系统进化树分析Figure 1. Phylogenetic tree analysis of the already identified microbial PLDs amino acid sequences
图 1 微生物PLD氨基酸序列的系统进化树分析Figure 1. Phylogenetic tree analysis of the already identified microbial PLDs amino acid sequences2.2 ADPLD蛋白序列的生物信息学分析
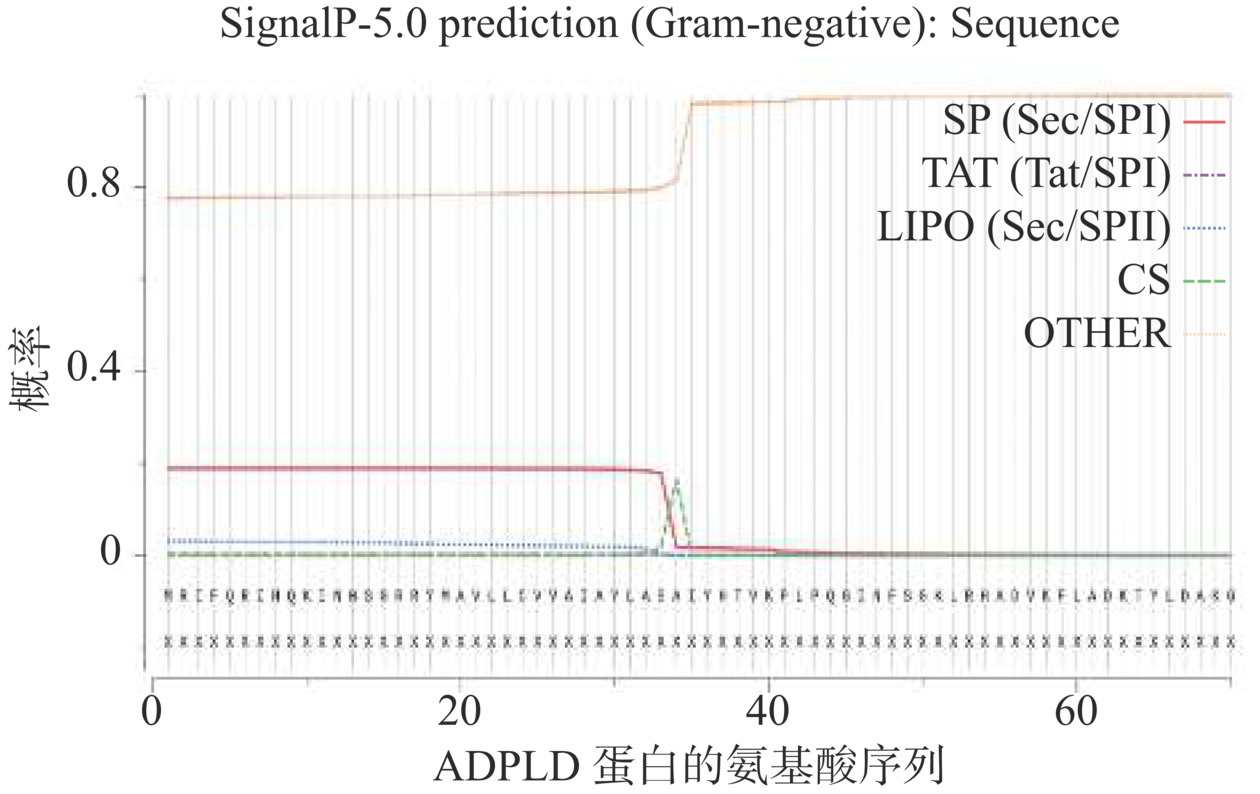
采用SignalP 5.0分析ADPLD全长基因的信号肽位置和切割位点。如图2所示,SP值在第1位氨基酸到第34位氨基酸的剪切位点之前较高,而在该剪切位点之后明显降低。因此可以判定ADPLD的信号肽切割位点在第34位丙氨酸和第35位异亮氨酸之间(Ala34-Ile35)。MRIFQRIHQKINWSGRRYMAVLLCVVAIAYLASA被预测为ADPLD的信号肽。选用大肠杆菌作为表达宿主时应保留第34位丙氨酸之后的成熟肽序列。ExPASY ProtParam分析表明,ADPLD的成熟肽含有453个氨基酸残基,分子量为51227.43 Da,蛋白不稳定系数(Instability index)为30.63,推测蛋白质稳定性强。ADPLD只有一个保守的HKD基序,分别位于His188、Lys190和Asp195。ADPLD的成熟肽属于疏水性蛋白质,总亲水性平均系数(GRAVY)为−0.368,带正电荷氨基酸残基(Arg,Lys)共有54个(占总氨基酸的11.9%),带负电荷氨基酸残基(Asp,Glu)共有49个(占总氨基酸的10.8%)。ADPLD理论等电点为8.79。
2.3 ADPLD基因的克隆
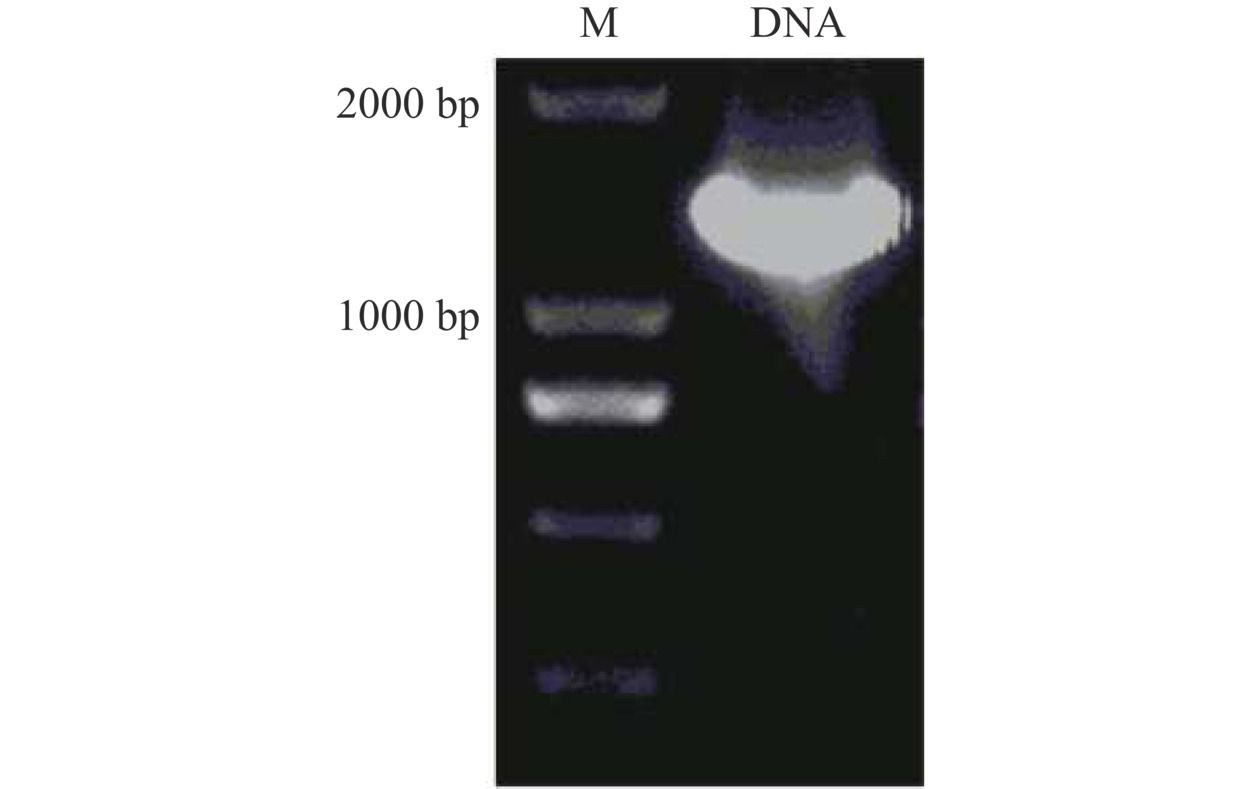
生物信息学分析表明ADPLD成熟肽含有453个氨基酸,一个密码子由3个碱基组成,因此ADPLD对应的基因序列长度为1359 bp。将ADPLD成熟肽编码的基因序列按照大肠杆菌密码子偏好性优化后,交由上海生工生物公司合成。ADPLD基因序列经过PCR扩增,酶切后与pET-28a载体连接。利用ADPLD基因上下游引物进行PCR反应,PCR产物的凝胶电泳结果如图3所示,从Acinetobacter sp. DUT-2中克隆得到的ADPLD成熟肽基因条带大小约为1300 bp,所示结果与ADPLD基因长度1359 bp基本一致。
![]() 图 3 PCR克隆ADPLD基因的凝胶电泳结果注:M,DL 2000 Marker;DNA,ADPLD基因。Figure 3. Gel electrophoresis result of ADPLD gene cloned by PCR
图 3 PCR克隆ADPLD基因的凝胶电泳结果注:M,DL 2000 Marker;DNA,ADPLD基因。Figure 3. Gel electrophoresis result of ADPLD gene cloned by PCR2.4 ADPLD酶蛋白的纯化与底物选择性分析
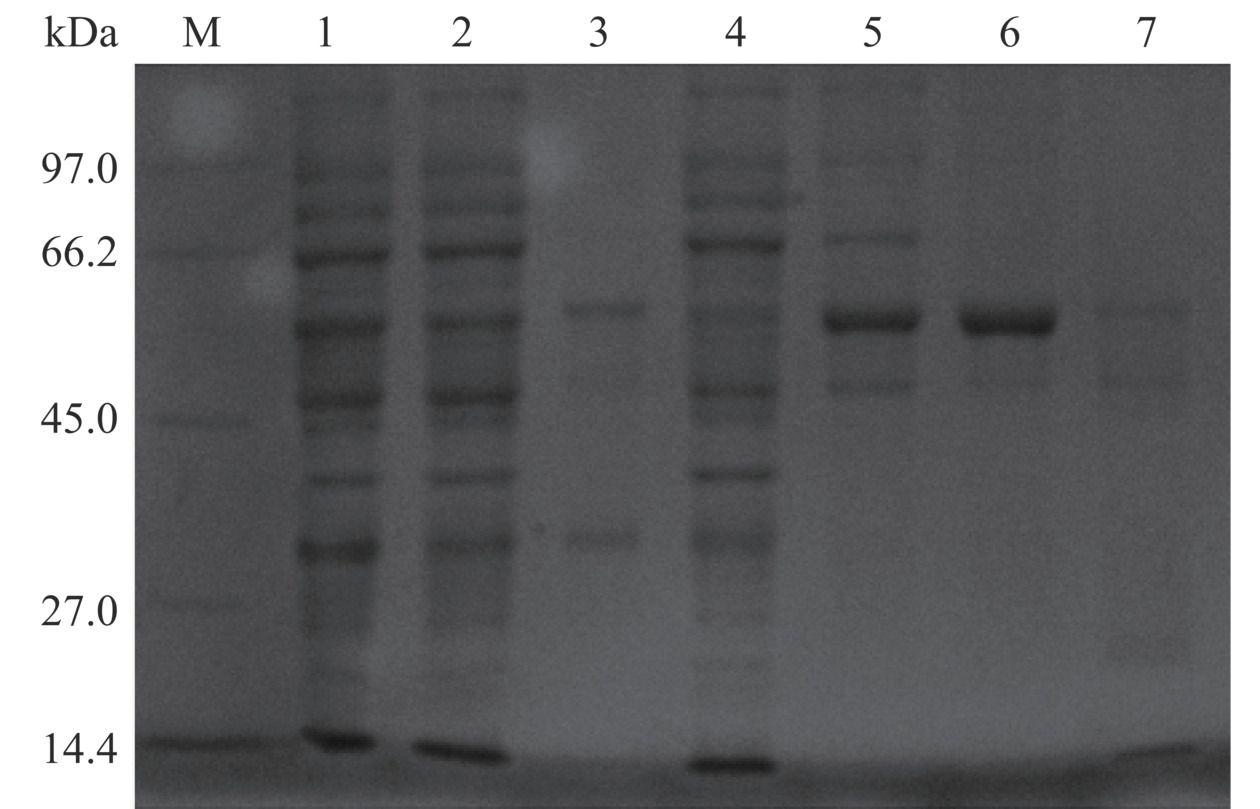
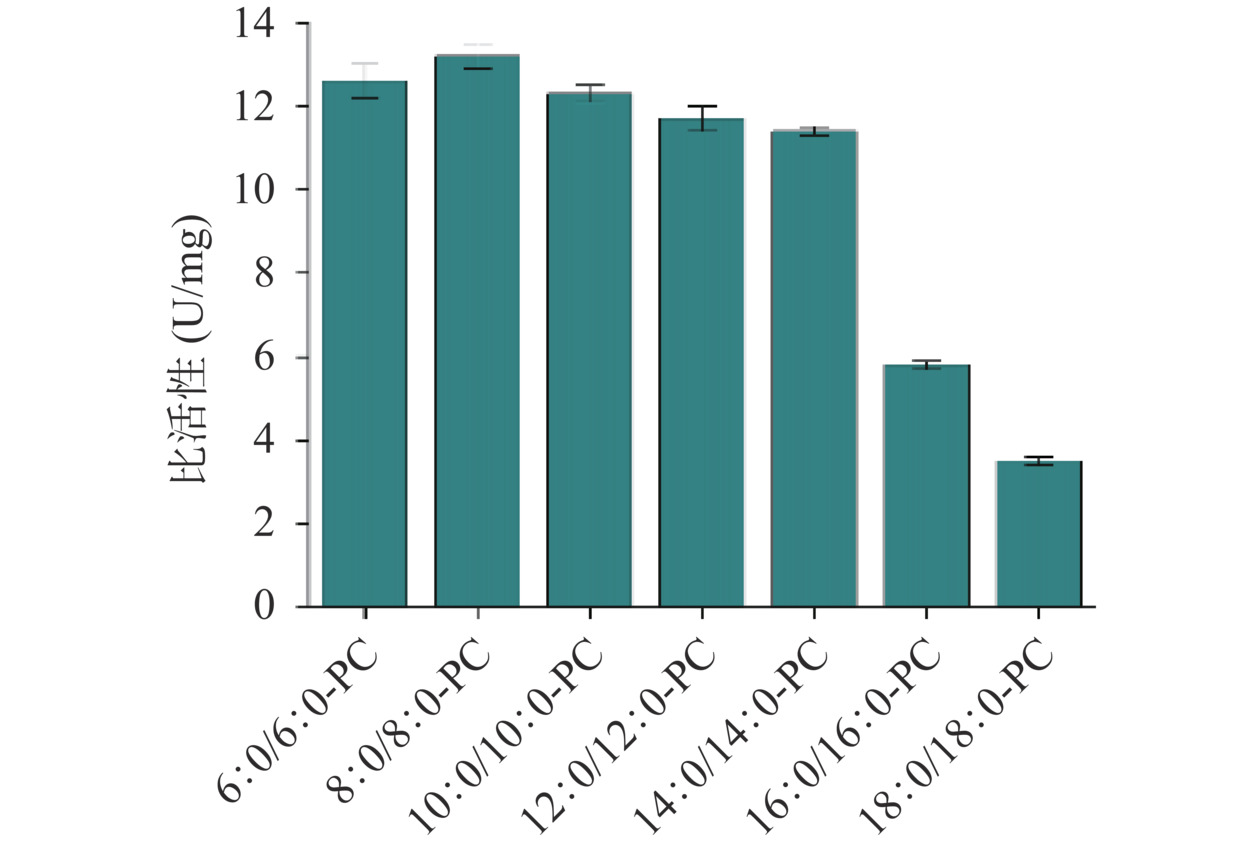
使用表达载体pET-28a和大肠杆菌Shuffle T7菌株能够成功获得ADPLD的可溶性蛋白。仅使用50 mmol/L咪唑浓度通过Ni2+亲和层析便能纯化出较为均一的蛋白(图4)。以大豆PC为天然底物时测定的ADPLD比活性约为4.09 U/mg。磷脂分子由一个极性的亲水头基和两个疏水的酰基链组成,亲水头基的类型和酰基链的长度影响磷脂分子的物理化学性质。目前研究集中在通过PLD介导的转磷脂酰化反应催化PC头部基团的转换来合成稀有磷脂,然而针对酰基链组成在PLD催化识别磷脂中的作用研究甚少。虽然PLD的催化位点是保守的,但几何形状高度可变的底物结合口袋为不同酰基链长的磷脂分子提供了弹性环境[38]。研究ADPLD对具有不同酰基链长PC底物的活性水平有利于提高对ADPLD催化的合成反应质量的认识。如图5所示,ADPLD对中和短链PC(C6~C14)的活性相对较高,其中ADPLD对8:0/8:0-PC的比活性最大,约为13.2 U/mg。当PC的酰基链长从C14增加至C16时,ADPLD对PC的活性出现明显下降。与ADPLD对8:0/8:0-PC的比活性相比,ADPLD对16:0/16:0-PC和18:0/18:0-PC的比活性分别下降了约56.06%和73.48%。目前仅有链霉菌来源的PLD对磷脂底物的酰基链长特异性被报道,研究指出S. antibioticus PLD对sn-2位为中和长酰基链的PC具有较高的催化活性,而对sn-2位为短酰基链的PC底物特异性较低[39]。综上所述,具有中和短酰基链的PC比长酰基链PC更适合作为ADPLD的底物,这一特性有利于在要求更宽底物范围的工业环境中对PLD介导的生物合成提供补充。
![]() 图 4 重组ADPLD表达与纯化电泳图注:M,中分子量蛋白Marker;泳道1,细胞总裂解液;泳道2,裂解上清液;泳道3,菌体沉淀;泳道4,穿过液;泳道5,Ni2+亲和层析中20 mmol/L咪唑洗脱样品;泳道6,Ni2+亲和层析中50 mmol/L咪唑洗脱样品;泳道7,Ni2+亲和层析中100 mmol/L咪唑洗脱样品。Figure 4. Electrophoresis of expression and purification of recombinant ADPLD
图 4 重组ADPLD表达与纯化电泳图注:M,中分子量蛋白Marker;泳道1,细胞总裂解液;泳道2,裂解上清液;泳道3,菌体沉淀;泳道4,穿过液;泳道5,Ni2+亲和层析中20 mmol/L咪唑洗脱样品;泳道6,Ni2+亲和层析中50 mmol/L咪唑洗脱样品;泳道7,Ni2+亲和层析中100 mmol/L咪唑洗脱样品。Figure 4. Electrophoresis of expression and purification of recombinant ADPLD![]() 图 5 纯化后的重组酶ADPLD对不同酰基链长PC的水解活力Figure 5. Hydrolysis activity of the purified recombinant enzyme ADPLD toward PCs with different acyl chain lengths
图 5 纯化后的重组酶ADPLD对不同酰基链长PC的水解活力Figure 5. Hydrolysis activity of the purified recombinant enzyme ADPLD toward PCs with different acyl chain lengths2.5 ADPLD底物识别的分子基础
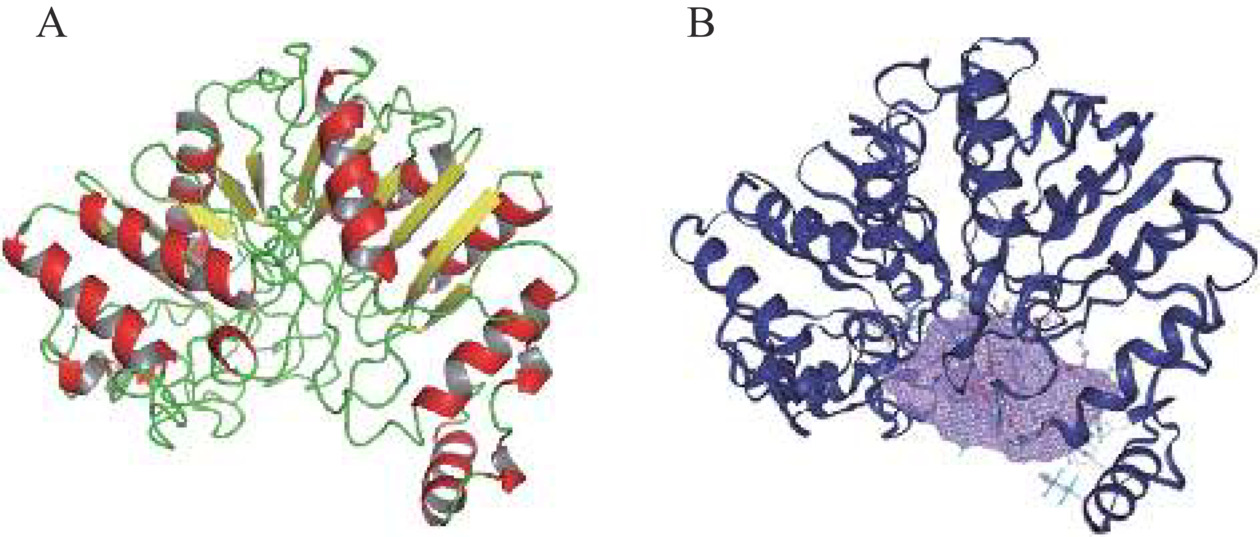
使用分子对接技术构建了Acinetobacter sp. DUT-2的PLD与配体(2R)-3-(phosphonooxy) propane-1,2-diyl diheptanoate(PDB ID:PD7)的复合物结构模型。图6A显示了ADPLD的三维模型结构,ADPLD包含13个α-Helix、15个β-Strand和部分Coil。此外,ADPLD底物结合口袋的面积为1174.44 Å2,空间体积为1131.52 Å3(图6B)。
![]() 图 6 ADPLD的三维结构模型注:红色结构为Helix,黄色结构为Strand,绿色部分为Coil。ADPLD底物结合口袋的体积形状显示为紫色网格。Figure 6. Three-dimensional (3D) structure model of ADPLD
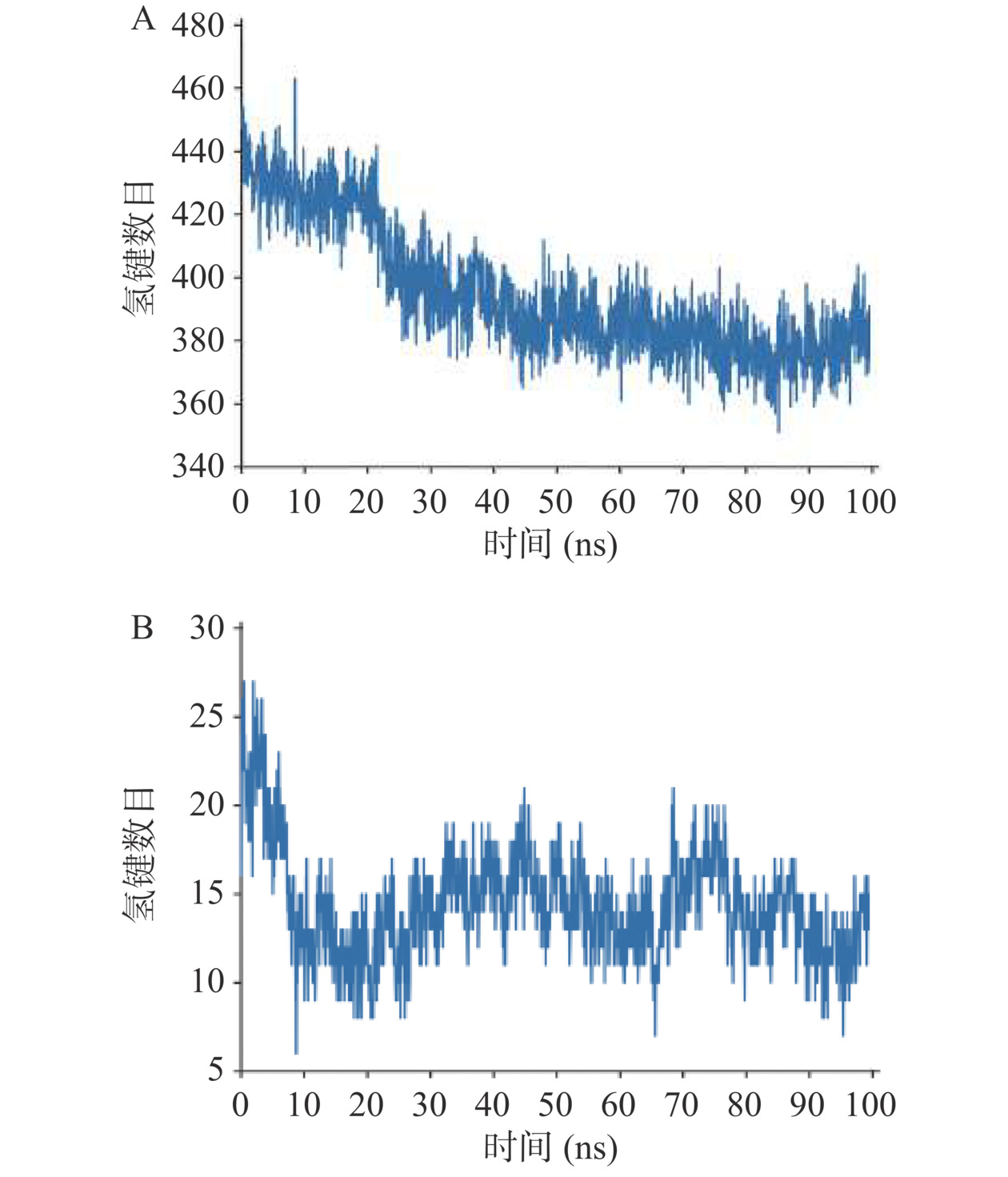
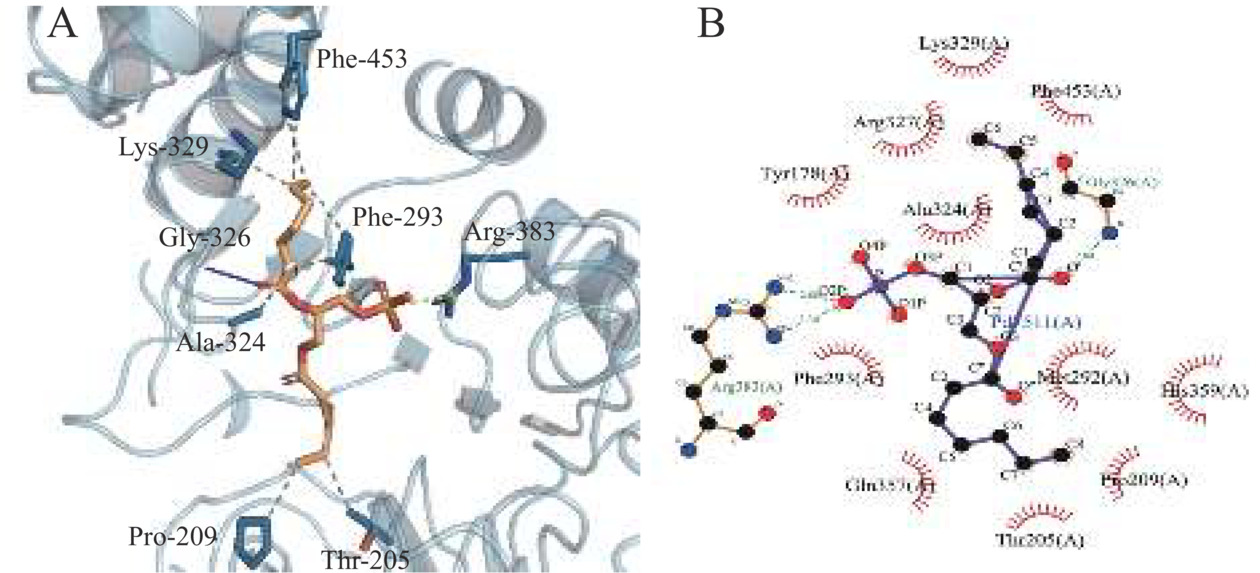
图 6 ADPLD的三维结构模型注:红色结构为Helix,黄色结构为Strand,绿色部分为Coil。ADPLD底物结合口袋的体积形状显示为紫色网格。Figure 6. Three-dimensional (3D) structure model of ADPLDPLD是一种在脂水界面完成催化反应的界面酶,构建了ADPLD在脂水界面的分子动力学模拟体系。从图7A中可以看出,ADPLD蛋白内氢键数目在50 ns模拟之后达到了平衡,约为380对氢键,对蛋白结构的稳定起主要作用。PLD的底物结合口袋与磷脂的非极性酰基链之间主要以疏水作用相结合[40]。平衡后,蛋白与周围磷脂分子间氢键数目约为12对(图7B),远小于蛋白内部氢键,推测ADPLD与磷脂分子间主要是疏水相互作用。因此进一步研究了当PC分子进入ADPLD底物结合口袋后受体与配体之间的相互作用(图8)。氢键和范德华作为分子间作用力有助于PC与ADPLD底物结合口袋紧密且稳定的结合。PLIP分析结果显示残基Thr205、Pro209、Phe293、Ala324、Lys329和Phe453能够分别与PC形成疏水相互作用。Arg383和Gly326能够与PC形成氢键,其中Arg383(N)、Gly326(N)与PC(P)之间的距离<3 Å。这些结果表明,ADPLD能够与磷脂分子形成一个稳定的酶与底物的中间体。
![]() 图 7 膜蛋白系统中ADPLD在100 ns分子动力学模拟中的氢键数量注:A.蛋白内的氢键数量变化;B.蛋白与PC单层膜间的氢键数量变化。Figure 7. Number of hydrogen bonds in 100 ns molecular dynamics simulation of ADPLD in membrane protein system
图 7 膜蛋白系统中ADPLD在100 ns分子动力学模拟中的氢键数量注:A.蛋白内的氢键数量变化;B.蛋白与PC单层膜间的氢键数量变化。Figure 7. Number of hydrogen bonds in 100 ns molecular dynamics simulation of ADPLD in membrane protein system![]() 图 8 (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate(PDB ID:PD7)配体与ADPLD的分子对接结果注:PC分子与ADPLD底物结合口袋的关键残基相互作用。在3D模型中,ADPLD底物结合口袋中与PC底物形成氢键和疏水相互作用的氨基酸显示为棒状结构,疏水相互作用展示为灰色虚线,氢键展示为蓝色实线。在2D模型中,氢键和疏水作用分别展示为绿色虚线和红色虚线。Figure 8. Molecular docking results of (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate ligand model (PDB ID:PD7) into the substrate-binding pocket of ADPLD
图 8 (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate(PDB ID:PD7)配体与ADPLD的分子对接结果注:PC分子与ADPLD底物结合口袋的关键残基相互作用。在3D模型中,ADPLD底物结合口袋中与PC底物形成氢键和疏水相互作用的氨基酸显示为棒状结构,疏水相互作用展示为灰色虚线,氢键展示为蓝色实线。在2D模型中,氢键和疏水作用分别展示为绿色虚线和红色虚线。Figure 8. Molecular docking results of (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate ligand model (PDB ID:PD7) into the substrate-binding pocket of ADPLD3. 结论
本研究从NCBI数据库中挖掘了一个来源于Acinetobacter sp. DUT-2 的PLD(ADPLD),依据大肠杆菌的密码子偏好性设计编码ADPLD的基因序列,构建大肠杆菌表达载体并在大肠杆菌表达系统中进行异源表达,分离纯化重组蛋白。研究发现ADPLD的分子量约为51 kDa,且只有一个HKD基序,这表明ADPLD的催化机制可能与传统认知中需要两个HKD基序完成PLD催化过程的反应机制有所不同。当ADPLD以大豆PC为底物时的比活性为4.09 U/mg。ADPLD对中和短链PC(C6~C14)的活性相对较高,当PC的酰基链长从C14增加至C16时,ADPLD对PC的活性出现明显下降。分子模拟结果推测ADPLD与周围磷脂间主要是疏水相互作用。分子对接结果表明ADPLD能够与磷脂分子形成一个稳定的酶与底物的中间体。本研究不仅提供了一个高催化活性的微生物来源PLD,而且为理解ADPLD的结构功能关系提供了重要信息。
-
![]()
图 1 微生物PLD氨基酸序列的系统进化树分析
Figure 1. Phylogenetic tree analysis of the already identified microbial PLDs amino acid sequences
![]()
图 3 PCR克隆ADPLD基因的凝胶电泳结果
注:M,DL 2000 Marker;DNA,ADPLD基因。
Figure 3. Gel electrophoresis result of ADPLD gene cloned by PCR
![]()
图 4 重组ADPLD表达与纯化电泳图
注:M,中分子量蛋白Marker;泳道1,细胞总裂解液;泳道2,裂解上清液;泳道3,菌体沉淀;泳道4,穿过液;泳道5,Ni2+亲和层析中20 mmol/L咪唑洗脱样品;泳道6,Ni2+亲和层析中50 mmol/L咪唑洗脱样品;泳道7,Ni2+亲和层析中100 mmol/L咪唑洗脱样品。
Figure 4. Electrophoresis of expression and purification of recombinant ADPLD
![]()
图 5 纯化后的重组酶ADPLD对不同酰基链长PC的水解活力
Figure 5. Hydrolysis activity of the purified recombinant enzyme ADPLD toward PCs with different acyl chain lengths
![]()
图 6 ADPLD的三维结构模型
注:红色结构为Helix,黄色结构为Strand,绿色部分为Coil。ADPLD底物结合口袋的体积形状显示为紫色网格。
Figure 6. Three-dimensional (3D) structure model of ADPLD
![]()
图 7 膜蛋白系统中ADPLD在100 ns分子动力学模拟中的氢键数量
注:A.蛋白内的氢键数量变化;B.蛋白与PC单层膜间的氢键数量变化。
Figure 7. Number of hydrogen bonds in 100 ns molecular dynamics simulation of ADPLD in membrane protein system
![]()
图 8 (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate(PDB ID:PD7)配体与ADPLD的分子对接结果
注:PC分子与ADPLD底物结合口袋的关键残基相互作用。在3D模型中,ADPLD底物结合口袋中与PC底物形成氢键和疏水相互作用的氨基酸显示为棒状结构,疏水相互作用展示为灰色虚线,氢键展示为蓝色实线。在2D模型中,氢键和疏水作用分别展示为绿色虚线和红色虚线。
Figure 8. Molecular docking results of (2R)-3-(phosphonooxy) propane-1,2-diyldiheptanoate ligand model (PDB ID:PD7) into the substrate-binding pocket of ADPLD
-
[1] CERMINATI S, PAOLETTI L, AGUIRRE A, et al. Industrial uses of phospholipases:current state and future applications[J]. Applied Microbiology & Biotechnology,2019,103(6):2571−2582.
[2] 袁雪丹, 王溪, 毛宗福, 等. 健康中国战略下国民健康与经济增长的联动关系研究-基于人力, 技术和产业视角[J]. 华东经济管理,2022,36(5):89−95. [YUAN X D, WANG X, MAO Z F, et al. Research on the linkage between national health and economic growth under the healthy China strategy:From the perspective of human capital, technology, and industry[J]. East China Economic Management,2022,36(5):89−95.] YUAN X D, WANG X, MAO Z F, et al. Research on the linkage between national health and economic growth under the healthy China strategy: From the perspective of human capital, technology, and industry[J]. East China Economic Management, 2022, 36(5): 89−95.
[3] ALI A H, ZOU X Q, ABED S M, et al. Natural phospholipids:Occurrence, biosynthesis, separation, identification, and beneficial health aspects[J]. Critical Reviews in Food Science & Nutrition,2019,59(2):253−275.
[4] 刘琦, 张海洋, 李雪晗, 等. 产磷脂酶D工程菌构建及发酵条件优化[J]. 食品研究与开发,2022,43(11):26−34. [LIU Q, ZHANG H Y, LI X H, et al. Engineering phospholipase D-producing strains of Bacillus subtilis and optimization of fermentation conditions[J]. Food Research and Development,2022,43(11):26−34.] doi: 10.12161/j.issn.1005-6521.2022.11.004 LIU Q, ZHANG H Y, LI X H, et al. Engineering phospholipase D-producing strains of Bacillus subtilis and optimization of fermentation conditions[J]. Food Research and Development, 2022, 43(11): 26−34. doi: 10.12161/j.issn.1005-6521.2022.11.004
[5] 周彦峰, 张涛, 江波, 等. 磷脂酰丝氨酸的酶法制备与分离研究进展[J]. 中国油脂,2018,43(10):53−57. [ZHOU Y F, ZHANG T, JIANG B, et al. Advance in enzymatic preparation and separation of phosphatidylserine[J]. China Oils and Fats,2018,43(10):53−57.] doi: 10.3969/j.issn.1003-7969.2018.10.011 ZHOU Y F, ZHANG T, JIANG B, et al. Advance in enzymatic preparation and separation of phosphatidylserine[J]. China Oils and Fats, 2018, 43(10): 53−57. doi: 10.3969/j.issn.1003-7969.2018.10.011
[6] SHIGEYUKI I, YOSHIFUMI H. Purification of Streptomyces chromofuscus phospholipase D by hydrophobic affinity chromatography on palmitoyl cellulose[J]. The Journal of Biochemistry,1979,85(1):79−95. doi: 10.1093/oxfordjournals.jbchem.a132334
[7] ZHOU W B, GONG J S, HOU H J, et al. Mining of a phospholipase D and its application in enzymatic preparation of phosphatidylserine[J]. Bioengineered,2018,9(1):80−89. doi: 10.1080/21655979.2017.1308992
[8] CHOOJIT S, BORNSCHEUER U T, UPAICHIT A, et al. Efficient phosphatidylserine synthesis by a phospholipase D from Streptomyces sp. SC734 isolated from soil-contaminated palm oil[J]. European Journal of Lipid Science and Technology,2016,118(5):803−813. doi: 10.1002/ejlt.201500227
[9] ZHAO Y, XU Y F, YU F, et al. Identification of a novel phospholipase D gene and effects of carbon sources on its expression in Bacillus cereus ZY12[J]. Journal of Microbiology,2018,56(4):264−271. doi: 10.1007/s12275-018-7529-1
[10] ZHANG Y N, LU F P, CHEN G Q, et al. Expression, purification, and characterization of phosphatidylserine synthase from Escherichia coli K12 in Bacillus subtilis[J]. Journal of Agricultural and Food Chemistry,2009,57(1):122−126. doi: 10.1021/jf802664u
[11] MAO X Z, LIU Q Q, QIU Y Q, et al. Identification of a novel phospholipase D with high transphosphatidylation activity and its application in synthesis of phosphatidylserine and DHA-phosphatidylserine[J]. Journal of Biotechnology,2017,249:51−58. doi: 10.1016/j.jbiotec.2017.03.029
[12] HU F, WANG H, DUAN Z Q, et al. A novel phospholipase D constitutively secreted by Ochrobactrum sp. ASAG-PL1 capable of enzymatic synthesis of phosphatidylserine[J]. Biotechnology Letters,2013,35(8):1317−1321. doi: 10.1007/s10529-013-1207-5
[13] WILDERMAN P J, VASIL A I, JOHNSON Z, et al. Genetic and biochemical analyses of a eukaryotic-like phospholipase D of Pseudomonas aeruginosa suggest horizontal acquisition and a role for persistence in a chronic pulmonary infection model[J]. Molecular Microbiology,2010,39(2):291−303.
[14] PROCYK K J, KOVARIK P, GABAIN A V, et al. Salmonella typhimurium and lipopolysaccharide stimulate extracellularly regulated kinase activation in macrophages by a mechanism involving phosphatidylinositol 3-kinase and phospholipase D as novel intermediates[J]. Infection & Immunity,1999,67(3):1011−1017.
[15] HODGSON A L, BIRD P, NISBET I T. Cloning, nucleotide sequence, and expression in Escherichia coli of the phospholipase D gene from Corynebacterium pseudotuberculosis[J]. Journal of Bacteriology,1990,172(3):1256−1261. doi: 10.1128/jb.172.3.1256-1261.1990
[16] ZHANG Z, CHEN M, XU W, et al. Microbial phospholipase D:Identification, modification and application[J]. Trends in Food Science & Technology,2020,96:145−156.
[17] HU R K, CUI R G, TANG Q Y, et al. Enhancement of phospholipid binding and catalytic efficiency of Streptomyces klenkii phospholipase D by increasing hydrophobicity of the active site loop[J]. Journal of Agricultural and Food Chemistry,2021,69(37):11110−11120. doi: 10.1021/acs.jafc.1c04078
[18] WILKINS M R, GASTEIGER E, BAIROCH A, et al. Protein identification and analysis tools in the ExPASy server[J]. Methods Molecular Biology,1999,112:531−552.
[19] SAITOU N, NEI M. The neighbor-joining method:A new method for reconstructing phylogenetic trees[J]. Molecular Biology Evolution,1987,4(4):406−425.
[20] SUDHIR K, GLEN S, MICHAEL L, et al. MEGA X:Molecular evolutionary genetics analysis across computing platforms[J]. Molecular Biology and Evolution,2018,35(6):1547−1549. doi: 10.1093/molbev/msy096
[21] DIMITROV R A, GOULIAMOVA D E. Biological sequence comparison, molecular evolution and phylogenetics[J]. Biotechnology & Biotechnological Equipment,2012,26(1):209−217.
[22] HU R K, CUI R G, LAN D M, et al. Acyl chain specificity of marine Streptomyces klenkii phospholipase D and its application in enzymatic preparation of phosphatidylserine[J]. International Journal of Molecular Sciences,2021,22(19):10580. doi: 10.3390/ijms221910580
[23] ZHENG W, ZHANG C X, LI Y, et al. Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations[J]. Cell Reports Methods,2021,1(3):100014. doi: 10.1016/j.crmeth.2021.100014
[24] ZHANG C, FREDDOLINO P L, ZHANG Y. COFACTOR:improved protein function prediction by combining structure, sequence and protein-protein interaction information[J]. Nucleic Acids Research,2017,45(W1):W291−W299. doi: 10.1093/nar/gkx366
[25] YANG J, ZHANG Y. I-TASSER server:New development for protein structure and function predictions[J]. Nucleic Acids Research,2015,43(W1):W174−W181. doi: 10.1093/nar/gkv342
[26] LASKOWSKI. R A, MACARTHUR M W, MOSS D S, et al. PROCHECK:A program to check the stereochemical quality of protein structures[J]. Journal of Applied Crystallography,1993,26(Part 2):283−291.
[27] BENKERT P, BIASINI M, SCHWEDE T, et al. Toward the estimation of the absolute quality of individual protein structure models[J]. Bioinformatics (Oxford, England),2011,27(3):343−350.
[28] BERMAN H M, WESTBROOK J, FENG Z, et al. The protein data bank[J]. Nucleic Acids Research,2000,28(1):235−242. doi: 10.1093/nar/28.1.235
[29] MARTÍNEZ L, ANDRADE R, BIRGIN E G, et al. PACKMOL:A package for building initial configurations for molecular dynamics simulations[J]. Journal of Computational Chemistry,2009,30(13):2157−2164. doi: 10.1002/jcc.21224
[30] VANOMMESLAEGHE K, HATCHER E, ACHARYA C, et al. CHARMM general force field:A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields[J]. Journal of Computational Chemistry,2010,31(4):671−690. doi: 10.1002/jcc.21367
[31] DELANO W L. The pymol molecular graphics system[J]. Proteins-Structure Function and Bioinformatics,2002,30:442−454.
[32] LASKOWSKI R A, SWINDELLS M B. LigPlot+:Multiple ligand-protein interaction diagrams for drug discovery[J]. Journal of Chemical Information and Modeling,2011,51(10):2778−2786. doi: 10.1021/ci200227u
[33] VOLKAMER A, GRIEWEL A, GROMBACHER T, et al. Analyzing the topology of active sites:On the prediction of pockets and subpockets[J]. Journal of Chemical Information and Modeling,2010,50(11):2041−2052. doi: 10.1021/ci100241y
[34] VOLKAMER A, KUHN D, GROMBACHER T, et al. Combining global and local measures for structure-based druggability predictions[J]. Journal of Chemical Information and Modeling,2012,52(2):360−372. doi: 10.1021/ci200454v
[35] UESUGI Y, HATANAKA T. Phospholipase D mechanism using Streptomyces PLD[J]. Biochimica et Biophysica Acta (BBA)- Molecular and Cell Biology of Lipids,2009,1791(9):962−969. doi: 10.1016/j.bbalip.2009.01.020
[36] YANG H Y, ROBERTS M F. Cloning, overexpression, and characterization of a bacterial Ca2+/-dependent phospholipase D[J]. Protein Science,2002,11(12):2958−2968. doi: 10.1110/ps.0225302
[37] BHUSHAN A, MCNAMEE M G. Correlation of phospholipid structure with functional effects on the nicotinic acetylcholine receptor. A modulatory role for phosphatidic acid[J]. Biophysical Journal,1993,64(3):716−723. doi: 10.1016/S0006-3495(93)81431-0
[38] PLEISS J, FISCHER M, SCHMID R D. Anatomy of lipase binding sites:The scissile fatty acid binding site[J]. Chemistry and Physics of Lipids,1998,93(1-2):67−80. doi: 10.1016/S0009-3084(98)00030-9
[39] DAMNJANOVIĆ J, NAKANO H, IWASAKI Y. Acyl chain that matters:Introducing sn-2 acyl chain preference to a phospholipase D by protein engineering[J]. Protein Engineering Design and Selection,2019,32(1):1−11. doi: 10.1093/protein/gzz019
[40] HU R K, WU L X, CHENG Q P, et al. Structural variations and phospholipid binding characteristics of Streptomyces klenkii phospholipase D at the lipid-water interface[J]. LWT-Food Science and Technology,2023,179:114672. doi: 10.1016/j.lwt.2023.114672
-
其他相关附件
-
PDF格式
2023090014-13 47KB
-
 下载:
下载:







 下载:
下载:

计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
