


Effect and Process Conditions of Cold Plasma Combined withL-Glutamic Acid and Salt Stress on Germination and Enrichment ofγ-Aminobutyric Acid in Adzuki Bean
-
摘要: 为探究等离子体联合盐胁迫对红小豆萌发后γ-氨基丁酸(γ-Aminobutyric acid,GABA)含量的富集作用及效果。本实验以红小豆为原料,考察大气冷等离子电压、频率、时间处理种子对其发芽过程中GABA含量的影响,同时采用L-谷氨酸(L-Glu)联合盐胁迫的发芽方法,通过考察单因素(发芽时间、CaCl2、L-Glu和NaCl浓度)对GABA富集量的影响及响应面优化试验确定该法富集GABA最佳工艺。结果表明,大气冷等离子体技术处理种子对其萌发富集γ-氨基丁酸有促进作用,电压90 kV、频率120 Hz、时间20 min条件下大气冷等离子体处理效果较好。在发芽时间为58 h、CaCl2浓度为4.4 mmol/L、L-Glu浓度为3.2 mg/mL、NaCl浓度为66 mmol/L时,发芽红小豆GABA含量为160.23±2.91 mg/100 g,是未发芽红小豆的7.12倍。该方法高效可靠且成本低,为富含GABA食品的工厂化生产提供技术参考。Abstract: This study aimed to investigate the effect of cold atmospheric pressure plasma (CAPP) treatment combined with salt stress on the enrichment of gamma-aminobutyric acid (GABA) in adzuki beans after germination. The effect of CAPP voltage, frequency, and duration of treatment of seeds, on their GABA content during germination was investigated using adzuki beans as raw material. In addition, the method of germination using L-glutamic acid (L-Glu) combined with salt stress was used to investigate the effect of single factors (germination time and concentrations of CaCl2, L-Glu, and NaCl) on enrichment of GABA. The optimal process conditions for enrichment of GABA using response surface optimization experiments were also determined. The results showed that the treatment of seeds with CAPP technology had a beneficial effect on their germination and enrichment of GABA. The CAPP treatment was more effective under the following conditions: voltage of 90 kV, frequency of 120 Hz, and duration of 20 min. When the germination time was 58 h and the CaCl2, L-Glu, and NaCl concentrations were 4.4 mmol/L, 3.2 mg/mL, and 66 mmol/L, respectively, the GABA content of germinated adzuki beans was 160.23±2.91 mg/100 g, which was 7.12 times higher than that of ungerminated adzuki beans. This method is efficient, reliable, cost-effective, and provides a technical reference for the industrial production of GABA-rich foods.
-
Keywords:
- cold plasma /
- L-glutamic acid /
- salt stress /
- adzuki beans /
- γ-aminobutyric acid
-
红小豆是一种高蛋白、低脂肪、多营养的小杂粮,在国际上有“红珍珠”的美称。γ-氨基丁酸(γ-Aminobutyric acid,GABA)是一种非蛋白质氨基酸,在动植物中微量存在但具有不可替代的作用。作为植物中一种逆环境生长因子,GABA在植物萌发生长阶段大量富集。GABA在动物体内是一种重要的抑制性神经递质,除此之外,还有安神、保肝护肾、降糖降压等多种生理功效。GABA在2009年被中国列为新资源食品,并规定其摄入量不得超过500 mg/d[1]。因此,利用植物富集法开发GABA相关功能性食品更容易得到消费者青睐。
豆类的发芽是一种植物凝集素积累,改善营养均衡的最廉价处理方式。发芽可以使豆子中大分子物质如淀粉、蛋白质和脂肪分解为小分子,还使其中具有生物功能的活性成分如GABA、多酚等急剧升高。国内外学者研究发现以豆类为原料,提供其发芽所需的一般或特殊环境条件,使其抗逆环境基因表达,影响初级代谢、次级代谢物合成,培养成营养价值更高的豆芽[2]。周一峰等[3]研究正常发芽条件下花豆发芽过程中GABA的变化,发现30 ℃浸泡14 h,35 ℃发芽22 h,GABA的含量最高为0.40 mg/g。左娜等[4]研究发现通过谷氨酸钠、抗坏血酸等诱导黄豆和绿豆发芽,GABA富集量分别达到0.3025 mg/g和0.2783 mg/g。Jiang等[5]研究发现,真空联合谷氨酸钠胁迫发芽使红小豆中的GABA含量显著提高,并对相关基因的表达研究发现谷氨酸脱羧酶(Glutamic acid decarboxylase,GAD)基因表达与GABA的积累呈正相关。吕秋洁等[6]研究比较了L-谷氨酸(L-Glutamic acid,L-Glu)和谷氨酸钠(Monosodium glutamate,MSG)诱导紫糙米发芽,发现L-Glu较MSG更利于GABA的积累。曾晴等[7]研究不同盐种类对大豆发芽富集GABA的影响,发现氯化钠和氯化钙胁迫发芽GABA含量明显高于氯化钾、氯化镁、氯化铁等其他盐类。因此,通过低氧、超声[8]、冻融[9]、化学物质诱导等技术方法生产高GABA豆芽或者其他食品是可行的且潜力巨大,尤其是在低温大气压等离子体[10]、酸性电生功能水[11]等新兴技术诱导粮食种子发芽生产高GABA食品方面缺乏探索性研究。
冷等离子体(Cold Plasma,CP)被认为是物质的第四种状态,仅次于固态、液态和气态。物理上,它是部分或完全电离的气体混合物,含有活性物质,如激发态粒子、活性氧(Reactive oxygen species,ROS)、活性氮(Reactive nitrogen species,RNS)和OH自由基[12]。现代研究发现等离子体和种子发芽之间的相互作用包括种皮改性、反应物种、种子灭菌、热量和紫外线辐射与分子现象(包括转录和表观遗传调控)相关的表达,可以促进生物活性物质积累,提高芽的生物活性[13−14]。因此使用CP提高豆芽中生物活性成分是芽菜行业可持续发展的一项有前途的技术。与热加工技术相比,冷等离子体技术具有节能环保、加工效率高、成本低、适用于大型系统等特点[15]。Chen等[16]研究了冷等离子体处理对发芽糙米中GABA含量变化的影响,糙米经冷等离子体处理(3000 kV,10 min)后发芽,其GABA的含量达到最高为28 mg/100 g。CP技术产品种类有很多,其中在大气压力下产生的非热等离子体,称之冷大气压力等离子体(Cold atmospheric pressure plasma,CAPP)[17]。考虑到未来工厂化生产的简便性及可行性,CAPP的优点在于不使用低压设备和通入惰性气体的方法下也可产生等离子体气体,因此CAPP更适用于高GABA芽菜工厂化生产的实际情况。
目前,CP已被证明在提高谷物GABA积累方面具有潜在的作用,但对发芽豆类的研究较少。因此,本实验旨在探究CAPP处理对红小豆发芽期GABA的影响,为冷等离子体技术在提高谷物活性物质应用提供研究基础。利用响应面试验对CAPP处理的红小豆发芽培养液进行优化,以期为高GABA芽菜生产提供一些理论参考。
1. 材料与方法
1.1 材料与仪器
红小豆 品种珍珠红,大小均匀,直径约0.5 cm,大庆农贸市场购置;γ-氨基丁酸(≥99.9%) Sigma公司;次氯酸钠、苯酚、乙醇、硼酸 分析纯,上海陆都化学试剂厂。
HH-2数显恒温水浴锅 金坛市科兴仪器厂;CPX3800H-C超声波清洗机 EMERSON公司;TU-1810PC紫外可见分光光度计 北京普析通用仪器有限责任公司;LPWS恒温恒湿培养箱 上海陆谱科技有限公司;CPS-1型大气压冷等离子体杀菌机 南京农业大学协同苏州屹润食品科技有限公司及南京屹润等离子科技有限公司联合开发。
1.2 实验方法
1.2.1 红小豆处理及萌发工艺流程
红小豆种子→等离子体处理→培养液浸泡→培养箱发芽→测定芽豆GABA
1.2.2 冷等离子体处理
将随机选取的红小豆种子(每组约200个)置于样品盒中平铺成一层,将样品盒塑封。打开箱门,将样品盒放在移动平台上缓慢上升至极板距离为40 mm处,设置冷等离子处理条件为:电压90 kV,频率120 Hz,反应时间20 min(放电时间60 s/次,放电次数20次)。关闭箱门,开始反应。
1.2.3 培养液浸泡
将冷等离子体处理的红小豆清水洗5遍并除去悬浮瘪粒,然后用0.1%的次氯酸钠浸泡20 min,用蒸馏水冲洗3遍,分别用CaCl2溶液、NaCl溶液、L-Glu溶液和蒸馏水浸泡15 h。
1.2.4 培养箱发芽
将浸泡后的红小豆放入发芽盒,盖上4层纱布,放入温度35 ℃湿度85%的培养箱中培养,平均每2 h向培养箱中红小豆喷淋1次相应溶液。
1.2.5 单因素实验设计
1.2.5.1 冷等离子体处理时间对GABA含量的影响
控制等离子体电压90 kV,频率120 Hz,设置反应时间为:0 min(放电时间60 s/次,放电次数0次)、10 min(放电时间60 s/次,放电次数10次)、20 min(放电时间60 s/次,放电次数20次)、30 min(放电时间60 s/次,放电次数30次)。使用蒸馏水培养液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.5.2 冷等离子体处理电压对GABA含量的影响
控制等离子体频率120 Hz,反应时间20 min,设置电压为:30、60、90、120 kV。使用蒸馏水培养液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.5.3 冷等离子体处理频率对GABA含量的影响
控制等离子体电压90 kV,反应时间20 min,设置频率为:60、90、120、150 Hz。使用蒸馏水培养液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.5.4 NaCl浓度对GABA含量的影响
控制等离子体电压90 kV,频率120 Hz,反应时间20 min。培养液分别使用25、50、75 mmol/L的NaCl溶液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.5.5 CaCl2浓度对GABA含量的影响
控制等离子体电压90 kV,频率120 Hz,反应时间20 min。培养液分别使用2、4、6 mmol/L的CaCl2溶液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.5.6 L-Glu浓度对GABA含量的影响
控制等离子体电压90 kV,频率120 Hz,反应时间20 min。培养液分别使用1、2、3 mg/mL的L-Glu溶液浸泡和喷淋,放入培养箱中发芽,每12 h取一次样,测定红小豆GABA含量。
1.2.6 红小豆芽培养液优化设计
通过对单因素实验结果的分析,按照Box-Behnken试验设计方案,以GABA含量为响应值,对发芽时间、CaCl2浓度、L-Glu浓度、NaCl浓度四个因素分别取三个水平进行响应面试验,通过Design Expert 13软件对29组试验数据进行分析,并预测等离子体处理富集γ-氨基丁酸的最佳工艺条件。试验因素及水平如表1所示。
表 1 Box-Behnken试验因素及水平Table 1. Factors and levels of Box-Behnken experiment水平 因素 A 发芽时间
(h)B CaCl2浓度
(mmol/L)C L-Glu浓度
(mg/mL)D NaCl浓度
(mmol/L)−1 36 2 1 25 0 48 4 2 50 1 60 6 3 75 1.2.7 GABA含量测定
参照姜秀杰等[18]的方法稍作修改,将样品烘干粉碎,过100目筛。用蒸馏水定容到50 mL(样品为1.0 g),30 ℃超声提取2 h,滤纸过滤。8000 r/min离心10 min,取上清液备用。将GABA标品配制成0.1、0.2、0.3、0.4、0.5 mg/mL的标准溶液绘制成标准曲线。取不同浓度标准液或样品上清液1 mL,加入1 mL硼酸缓冲溶液(pH9.0),2 mL 5%重篜酚溶液(w/v),1 mL 10%次氯酸钠溶液,充分混匀。沸水浴10 min,冰水浴降温约15 min,溶液颜色由黄色慢慢变为蓝绿色,待化合物颜色稳定后加入60%乙醇2 mL,再次混合均匀。645 nm波长下测定吸光度。
1.3 数据处理
所有实验结果均重复3次,使用Origin 2022软件作图和IBM SPSS Statistics 26软件对数据进行显著性分析,使用Design-Expert 13软件进行Box-Behnken试验设计及处理。
2. 结果与分析
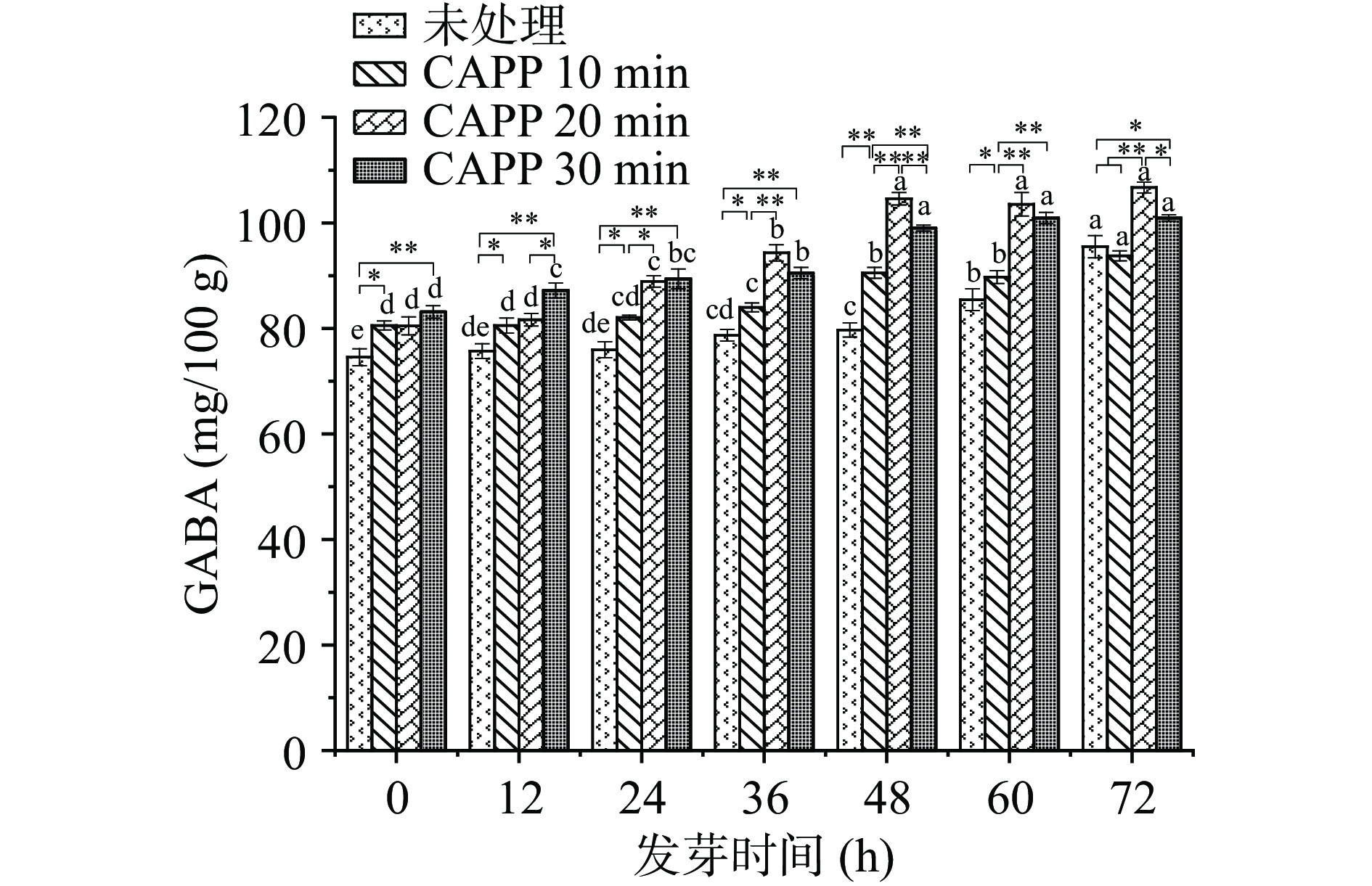
2.1 未处理与不同CAPP时间处理对红小豆发芽过程中GABA含量的影响
本实验中从种子吸水达到饱和的状态开始计时,即发芽0 h。在正常发芽情况下,发芽72 h红小豆的营养生长阶段基本结束并开始生殖生长阶段[19],此时其内部结构发生较大改变不再适合人类食用,因此发芽时间在72 h之后的芽豆不再测定GABA。
GABA属于抗逆环境的活性物质,起促进生长代谢作用,随发芽时间不断延长,在芽成熟后植物体内便也达到平衡状态[20−21]。观察未处理与处理组红小豆发芽过程中GABA富集量的变化见图1,在发芽0~72 h内,红小豆在正常发芽条件(未等离子体处理)下,随发芽时间的延长GABA含量不断增加;CAPP处理的红小豆随发芽时间的增加GABA含量不断增加,但在发芽48 h后不再明显升高。造成这种现象的原因是等离子体可以缩短种子萌发期,Filatova等[22]研究等离子体有促进种子早熟作用,结果表明石竹种子在等离子体作用下,种子胚内的生命物质被激活,种皮软化,从而加快了种子萌发和出苗的速度。
![]()
未处理与不同CAPP时间处理的红小豆在同一发芽时间GABA富集量的变化见图1。在发芽0~24 h阶段,正常发芽的红小豆GABA含量缓慢上升,而等离子体处理组GABA含量增加明显。造成该现象的原因可能是CAPP处理的红小豆吸水速度快,吸水程度高,快速激活了种子中的内源酶。种子吸收水分程度取决于三个因素:种子的组成、种皮的湿润性和水分[23]。相同种子和水分情况下,等离子体处理可以改变种子表面的化学结构和粗糙度,导致种子的湿润性发生巨大变化。已有研究证实空气等离子体处理后种皮与水的表观接触角急剧减小,导致种子的湿润性增加[24−25]。ALVES JUNIOR等[26]更深入研究等离子体处理对豆科种子吸水两大部位(珠孔和肺门)的变化,发现离子体处理增加肺门的水分吸收量,同时珠孔表现出更开放的形态。在发芽24~48 h时,CAPP处理20 min和30 min的红小豆GABA富集效果都较好。在发芽48~72 h时,只有CAPP处理20 min组的GABA富集效果较好,且含量较高。造成这种情况的原因可能是等离子体处理种子的时间越长,ROS和RNS对种子的刺激越大,伤害越深。因此,在红小豆发芽前期等离子体刺激越大合成GABA越多。当等离子体时间为20 min,在发芽48 h就达到顶峰,为104.601±1.192 mg/100 g。发芽72 h时最高为106.718±1.030 mg/100 g,但从成本方面考虑选择等离子体处理时间为20 min,发芽时间48 h。
2.2 CAPP电压对红小豆发芽过程中GABA含量的影响
控制冷等离子体装置的时间为20 min,频率为120 Hz不变,观察红小豆正常发芽与不同等离子体电压在发芽过程中GABA富集量的变化。由图2可知,使用不同电压等离子体处理的红小豆,其GABA含量随发芽时间延长而不断增加,最后趋于平衡。其中等离子体电压为120 kV处理红小豆,其GABA含量呈先上升后下降趋势。除电压为120 kV外,各组等离子处理的红小豆都呈上升趋势,且电压越大,红小豆GABA含量越高。原因可能是种子在等离子体处理过程中,电压越高空气被电离的混合物越多,混合物中多种活性基团和粒子能够与种皮及表面微生物的发生更强烈的反应[27],从而促进种子吸水,阻碍微生物生长,使种子更易发芽,GABA含量升高。若对红小豆离子体处理电压过高,则没有上述的生长代谢趋势,反而有部分的豆子表面出现明显的电流打击的伤痕,其伤口破坏了豆子的免疫屏障,容易被有害微生物入侵。当等离体电压为120 kV,红小豆发芽时间为24~72 h阶段内,其GABA呈先下降后不变的趋势,说明电压过大时对种子的萌发以及GABA富集有抑制作用,其抑制机理有待深入研究。何瑞等[28]研究也发现相同结果,当大气压等离子体处理电压为3.4 kV时最有利于穿心莲种子萌发,继续加大电压,当大气压等离子体处理电压为5.95 kV时则对穿心莲种子的萌发有强烈抑制作用。因此控制一个合适的等离子体电压才能最有利于种子萌发,针对不同种子其最佳等离子体电压也不同。因此,针对CAPP处理红小豆种子最佳电压为90 kV。
![]() 图 2 CAPP电压对红小豆发芽过程中GABA含量的影响Figure 2. Effect of CAPP voltage on GABA content during germination of adzuki bean
图 2 CAPP电压对红小豆发芽过程中GABA含量的影响Figure 2. Effect of CAPP voltage on GABA content during germination of adzuki bean2.3 CAPP频率对红小豆发芽过程中GABA含量的影响
控制冷等离子体装置的时间为20 min,电压为90 kV不变,观察不同等离子体频率条件下红小豆发芽过程中GABA富集量的变化。由图3可知,不同频率的等离子体处理红小豆,随萌发时间的延长,其GABA含量呈先升高后平稳的趋势。等离子体处理频率越高其GABA富集的含量也越高。120 Hz和150 Hz等离子体处理红小豆,发芽48 h之后GABA含量分别达到最高为104.601±1.192 mg/100 g和101.467±2.225 mg/100 g,两者无显著性差异,继续发芽其GABA含量也无明显变化。已有研究表明通过优化原料基质特性(如水分含量)和调整等离子体电源设置(电压,频率)以增加反应物质的产生,可以提高微生物灭活效率[29]。但因红小豆表面呈拓扑结构,具有空隙和波纹化的粗糙表面可以在一定程度上保护植物细胞和表面微生物免受等离子体产生的ROS和RNS侵害[30]。改变冷大气压等离子体机器的频率是加速种子吸水,发芽,增加植物幼苗生长的有效手段。频率小于90 Hz时等离子体的处理效率较低,对微生物抑制作用较低,对刺激红小豆发芽作用也较弱,频率在120~150 Hz时等离子体处理对红小豆的刺激较大,有利GABA的富集。因此,为延长机器使用寿命,等离子体处理红小豆的最佳频率为120 Hz。
![]() 图 3 CAPP频率对红小豆发芽过程中GABA含量的影响Figure 3. Effect of CAPP frequency on GABA content during germination of adzuki bean
图 3 CAPP频率对红小豆发芽过程中GABA含量的影响Figure 3. Effect of CAPP frequency on GABA content during germination of adzuki bean2.4 NaCl对红小豆发芽过程中GABA含量的影响
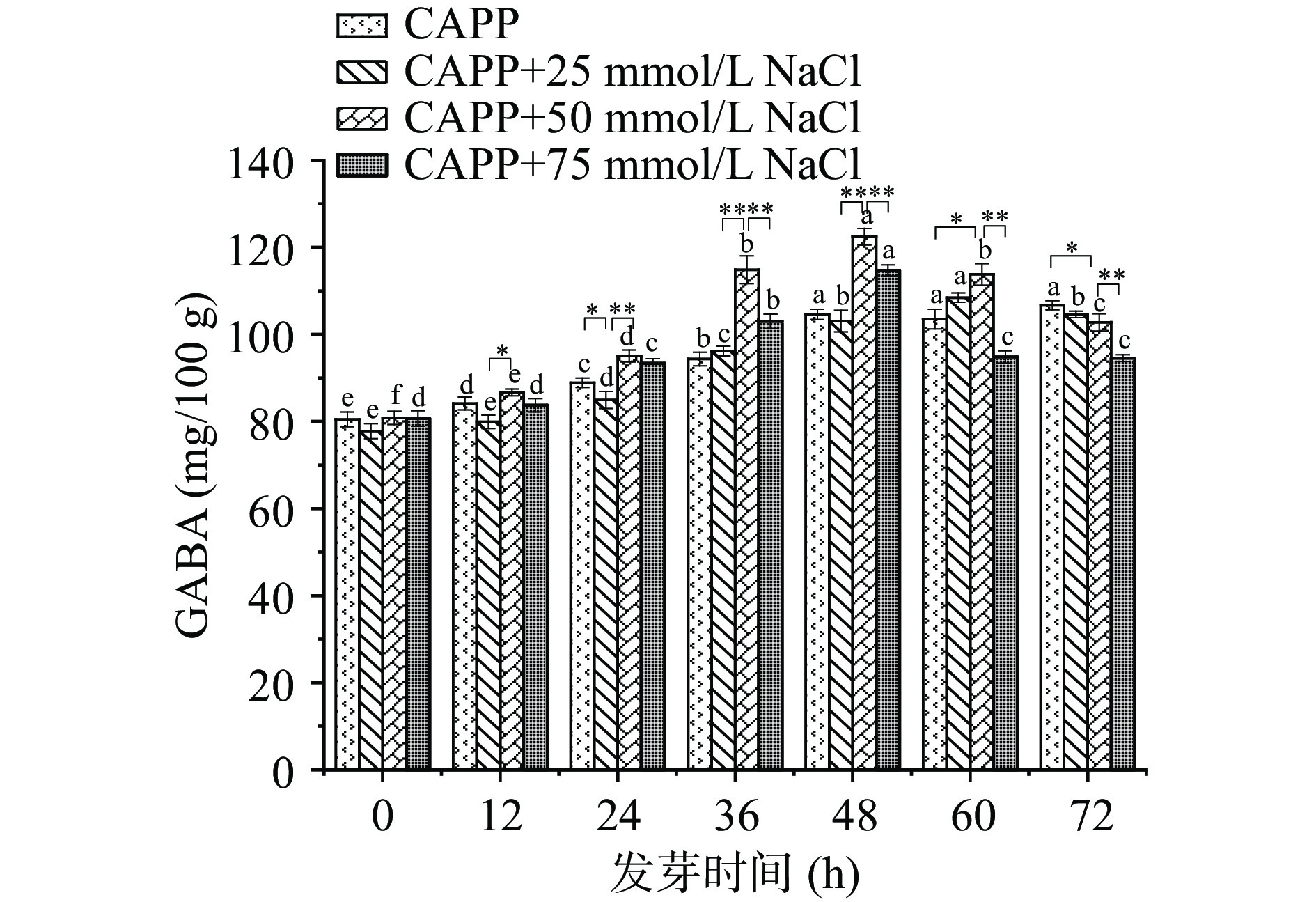
将等离子体处理的红小豆分为4组,依次喷洒不同浓度的NaCl溶液,观察不同浓度NaCl溶液处理的红小豆发芽过程中GABA的富集情况。由图4可知,CAPP+NaCl处理组的GABA含量高于等离子体处理组。随着NaCl的浓度增加,GABA的含量呈先升高后减小的趋势,当发芽时间为48 h,NaCl浓度为50 mmol/L时,GABA含量达到最高为122.456±1.888 mg/100 g。这是因为NaCl对植物富集GABA的两条途径都有促进效果。AL-QURAAN等[31]研究NaCl对发芽小麦进行处理发现5个品种小麦的GAD基因表达上调。因此,在GABA支路途径中NaCl会上调红小豆GAD的相关基因表达,提高GAD酶活性,富集GABA。在多胺降解途径中,XING等[32]研究表明在NaCl处理下,大豆胚根的二胺氧化酶(Diamine oxidase,DAO)活性增强,促进多胺(Polyamines,PAS)降解,富集GABA。Yang等[33]研究发现NaCl处理使豆芽氨基醛脱氢酶(Aminoaldehyde dehydrogenase,AMADH)、GAD和DAO活性分别提高了39.7%、28.4%和21.2%。除此之外,还有研究表明NaCl刺激植物产生氧化应激,不仅会提高超氧化物歧化酶和过氧化氢酶活性,还能增加苯丙氨酸氨裂解酶,肉桂酸4-羟化酶和4-香豆酸辅酶A连接酶的活性和表达,这些反应与表达也会减小GABA合成抑制剂的作用[34−35]。因此,25~75 mmol/L的NaCl培养液有利于CAPP红小豆富集GABA,50 mmol/L NaCl培养液效果最佳。
![]() 图 4 NaCl对红小豆发芽过程中GABA含量的影响Figure 4. Effect of NaCl on GABA content during germination of adzuki bean
图 4 NaCl对红小豆发芽过程中GABA含量的影响Figure 4. Effect of NaCl on GABA content during germination of adzuki bean2.5 CaCl2对红小豆发芽过程中GABA含量的影响
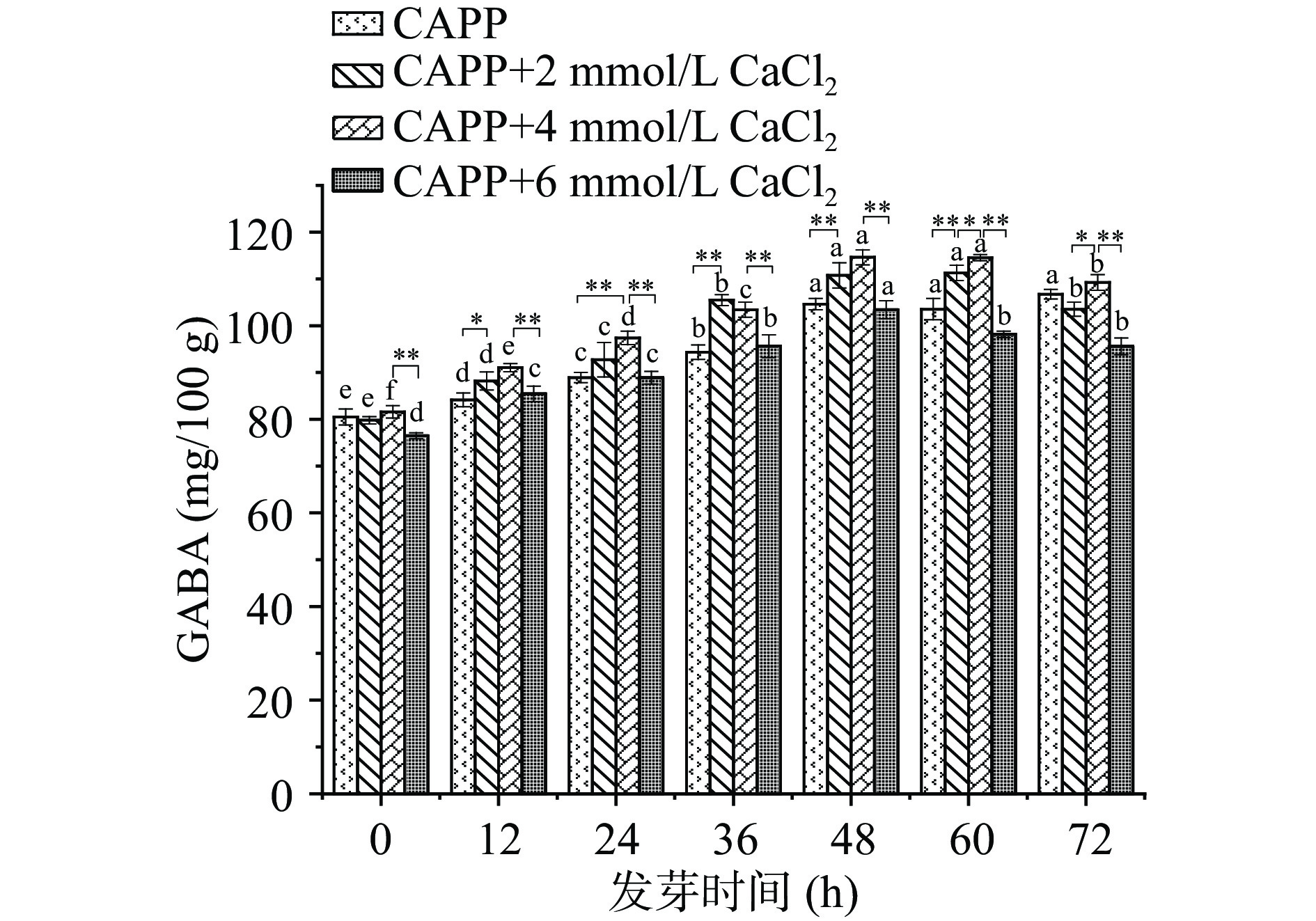
将正常发芽的红小豆分为4组,依次喷洒不同浓度的CaCl2溶液,观察不同浓度CaCl2溶液处理的红小豆发芽过程中GABA的富集情况。由图5可知,CAPP+CaCl2处理的豆子随发芽时间延长,GABA呈先升高后降低的趋势。2和4 mmol/L CaCl2处理的红小豆在发芽在48 h后才有明显差异,4 mmol/L CaCl2处理的红小豆富集效果更好。4 mmol/L CaCl2处理的红小豆在发芽48 h时达到最高为114.62±1.593 mg/100 g。植物中的GABA合成主要来自GABA支路中由谷氨酸脱羧酶(GAD)催化的不可逆的α-谷氨酸脱羧反应[36]。GAD是一种Ca2+/钙调蛋白依赖型酶,具有一个钙调蛋白结合区,其活性受Ca2+/Ca M控制[37]。王珊珊等[38]研究表明提高培养液中Ca2+浓度,促进Ca M区域与钙离子结合可以提高GAD的活性,从而加快谷氨酸合成GABA。朱云辉等[39]研究发现Ca2+浓度达到6.0 mmol/L时,发芽苦荞中GABA含量达到最大,此后随着Ca2+浓度增大,GABA含量呈减小趋势。因此,CaCl2在2~6 mmol/L范围内处理红小豆发芽有助于富集GABA,4 mmol/L的CaCl2培养液效果最佳。
![]() 图 5 CaCl2对红小豆发芽过程中GABA含量的影响Figure 5. Effect of CaCl2 on GABA content during germination of adzuki bean
图 5 CaCl2对红小豆发芽过程中GABA含量的影响Figure 5. Effect of CaCl2 on GABA content during germination of adzuki bean2.6 L-Glu对红小豆发芽过程中GABA含量的影响
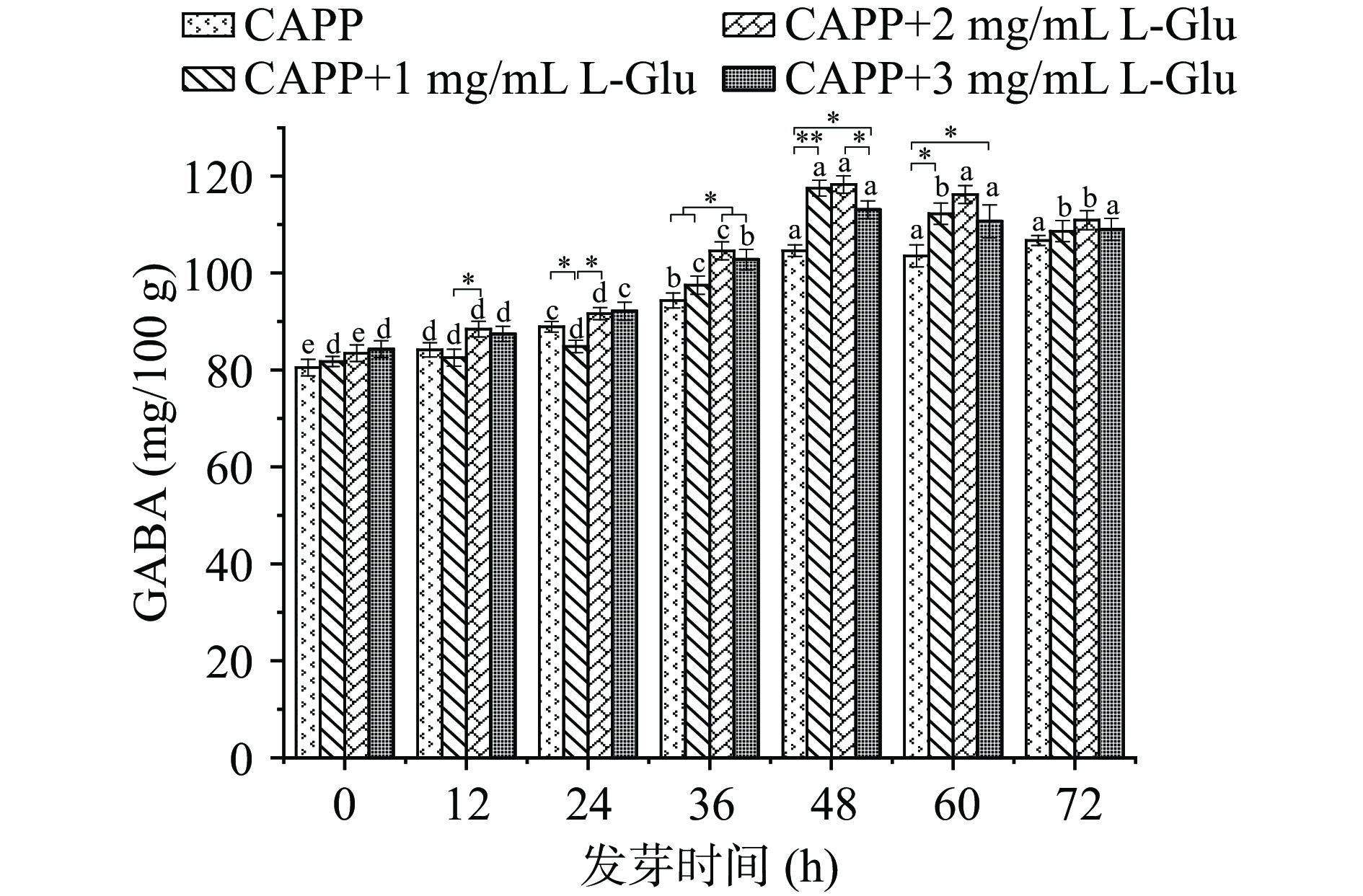
将正常发芽的红小豆分为4组,依次喷洒不同浓度的L-Glu溶液,观察不同浓度L-Glu溶液处理的CAPP红小豆发芽过程中GABA的富集情况。由图6可知,随发芽时间的延长,GABA的含量呈先增加后下降的趋势,当使用L-Glu浓度为1、2 mg/mL处理CAPP红小豆时,发芽48 h达到最大为117.518±1.634、118.264±1.804 mg/100 g。L-Glu是谷氨酸脱羧酶(GAD)的唯一底物[40]。在外源溶液中添加L-Glu可以提高植物的渗透调节能力,增加底物从而增加GABA含量。长时间限制谷氨酰胺合成,或减少蛋白质合成,或增加蛋白质降解,可使GABA富集,这是由于谷氨酸含量增多可使GABA支路中碳流量增加,并调节谷氨酸脱羧酶的活性[41]。陈慧等[42]用1.0 mg/mL的MSG溶液处理蚕豆发芽,结果发现GAD活性变化趋势与GABA含量变化相似。因此,使用1~3 mg/mL的L-Glu培养液可以提高红小豆GAD的酶活,增加GABA含量。2和3 mg/mL的L-Glu培养液效果都较好,且差别不大。
![]() 图 6 L-Glu对红小豆发芽过程中GABA含量的影响Figure 6. Effect of L-Glu on GABA content during germination of adzuki bean
图 6 L-Glu对红小豆发芽过程中GABA含量的影响Figure 6. Effect of L-Glu on GABA content during germination of adzuki bean2.7 响应面优化GABA富集工艺
2.7.1 回归模型的建立及方差分析
通过单因素实验结果,发现冷等离子体处理90 kV,120 Hz,20 min红小豆种子有利于发芽和富集GABA,且NaCl、CaCl2和L-Glu溶液三种诱导环境下,红小豆发芽后GABA的富集量都明显提高。确定了4个因素对L-Glu联合盐胁迫发芽CAPP红小豆富集GABA的影响的4水平,对29个试验组进行3次重复测量,响应面试验设计及结果见表2。
表 2 响应面试验设计及结果Table 2. Response surface design and results实验号 因素 GABA含量(mg/100 g) A B C D 1 1 1 0 0 146.92±5.08 2 1 0 0 -1 137.74±6.31 3 0 0 0 0 157.01±7.06 4 −1 0 0 1 114.76±1.09 5 0 −1 −1 0 130.07±2.78 6 0 0 0 0 160.34±2.09 7 0 0 0 0 159.32±4.88 8 0 0 0 0 152.48±3.38 9 −1 1 0 0 128.94±4.49 10 −1 0 −1 0 123.56±4.74 11 0 1 0 1 151.84±6.19 12 −1 −1 0 0 119.56±4.89 13 0 −1 1 0 120.30±3.97 14 1 0 1 0 139.03±5.92 15 0 0 1 −1 122.17±6.28 16 0 1 0 −1 125.91±4.86 17 1 0 0 1 166.11±6.38 18 0 0 −1 −1 133.12±6.96 19 0 −1 0 1 143.23±5.95 20 1 −1 0 0 141.22±3.39 21 0 −1 0 −1 136.52±4.18 22 −1 0 1 0 121.96±7.12 23 0 0 0 0 155.92±5.02 24 0 1 −1 0 142.95±5.25 25 −1 0 0 −1 122.77±3.29 26 1 0 −1 0 154.75±4.69 27 0 0 −1 1 136.78±2.94 28 0 1 1 0 137.11±5.86 29 0 0 1 1 157.37±7.75 在此基础上,利用Design-Expert 13软件对响应面进行设计与分析,并建立回归模型,得出二元回归方程为:
R1=157.01+12.85A+3.56B−1.94C+7.66D−0.9194AB−3.53AC+9.10AD+0.9840BC+4.80BD+7.89CD−11.95A2−11.05B2−11.72C2−8.08D2
回归模型方差分析结果见表3。由表3可知,一次项A、D与交互项AD和二次项A2、B2、C2、D2表现为高度显著(P<0.01),一次项B与交互项CD表现为显著(P<0.05)。模型总体极显著(P<0.0001),说明预测结果与真实结果有相关性,决定系数R2=0.8970,失拟项不显著(P˃0.05),说明残差是由随机误差引起。因此可知该模型拟合度较高,且误差在可接受范围内,表明可用于预测L-Glu联合盐胁迫发芽CAPP红小豆富集GABA工艺参数。根据二元回归方程中一次项系数绝对值的大小,判定各因素对响应值影响的主次顺序为发芽时间(A)˃NaCl浓度(D)˃CaCl2浓度(B)˃L-Glu浓度(C)。
表 3 响应面回归模型方差分析Table 3. Analysis of variance with response surface regression model来源 偏平方和 自由度 均方和 F值 P值 显著性 模型 5583.97 14 398.85 12.30 <0.0001 significant A 1981.87 1 1981.87 61.11 <0.0001 B 152.44 1 152.44 4.70 0.0479 C 45.27 1 45.27 1.40 0.2571 D 703.19 1 703.19 21.68 0.0004 AB 3.38 1 3.38 0.1043 0.7516 AC 49.86 1 49.86 1.54 0.2354 AD 330.96 1 330.96 10.21 0.0065 BC 3.87 1 3.87 0.1194 0.7348 BD 92.34 1 92.34 2.85 0.1137 CD 248.79 1 248.79 7.67 0.0150 A² 926.77 1 926.77 28.58 0.0001 B² 791.57 1 791.57 24.41 0.0002 C² 891.33 1 891.33 27.49 0.0001 D² 423.43 1 423.43 13.06 0.0028 残差 454.01 14 32.43 失拟项 415.86 10 41.59 4.36 0.0843 not significant 纯误差 38.15 4 9.54 总和 6037.97 28 2.7.2 响应面交互作用分析
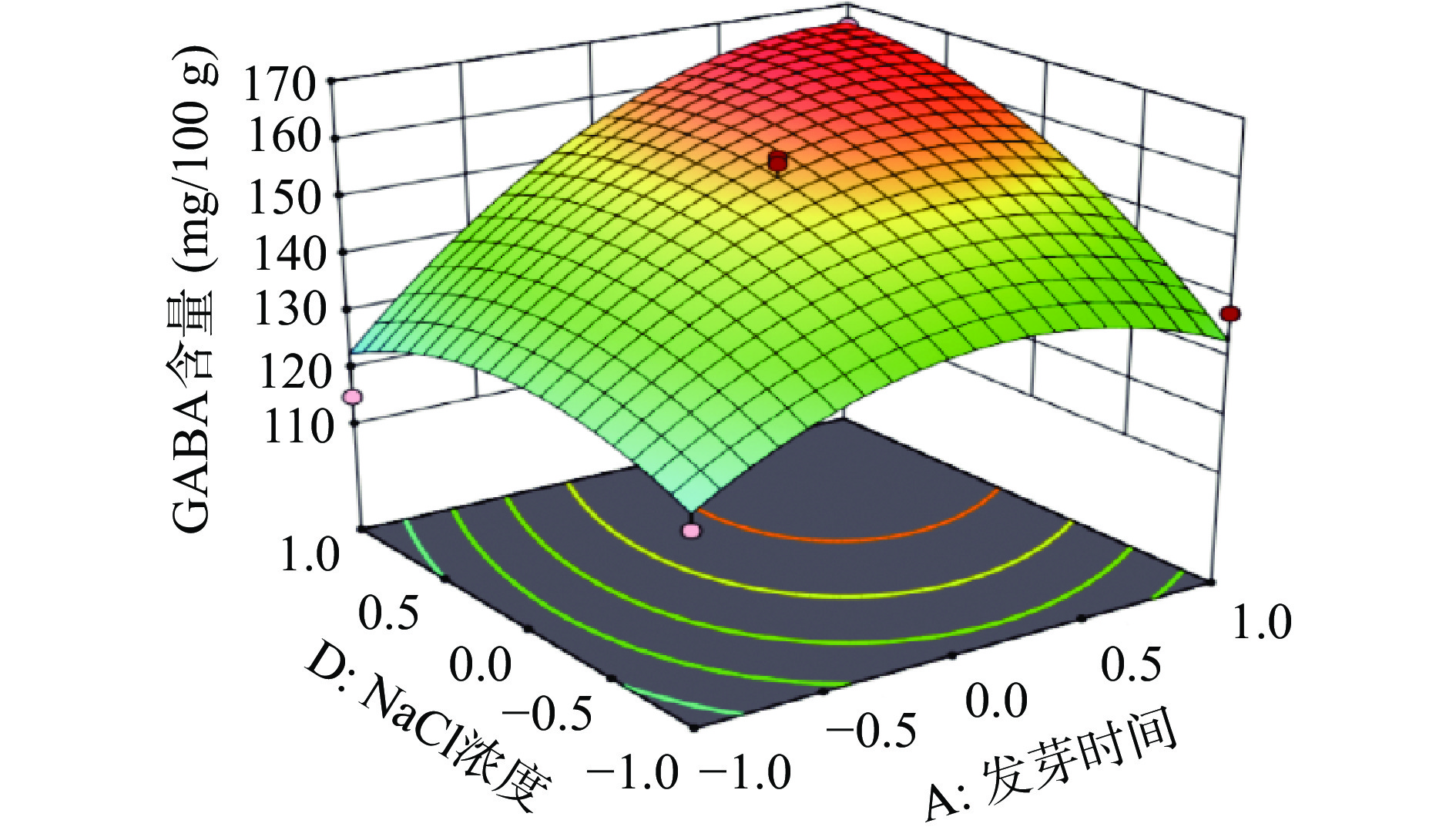
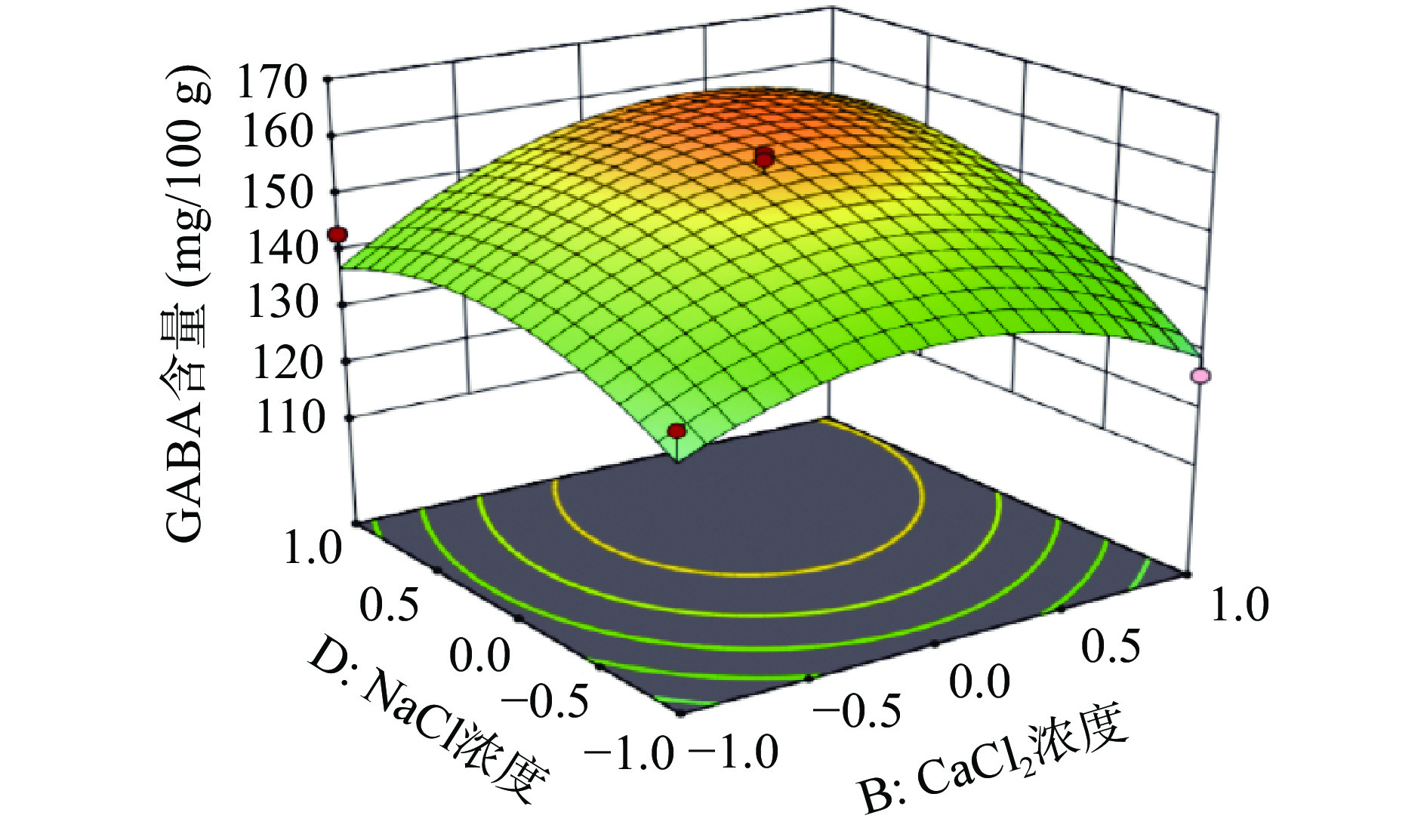
响应面是用于描述因素与响应值之间关系的三维空间曲面图,可直观反映出各因素之间的交互作用,及其对L-Glu联合盐胁迫发芽富集CAPP红小豆GABA含量的影响程度。图7~图8呈现的是对红小豆富集GABA有显著交互作用的响应面和等高线图。响应面坡度越陡峭,则表明响应值越敏感。
![]() 图 7 发芽时间和NaCl浓度交互作用的响应面图Figure 7. Response surface plots of the interaction between germination time and NaCl concentration
图 7 发芽时间和NaCl浓度交互作用的响应面图Figure 7. Response surface plots of the interaction between germination time and NaCl concentration![]() 图 8 CaCl2和NaCl浓度交互作用的响应面图Figure 8. Response surface plots of the interaction between CaCl2 and NaCl concentrations
图 8 CaCl2和NaCl浓度交互作用的响应面图Figure 8. Response surface plots of the interaction between CaCl2 and NaCl concentrations发芽时间、NaCl浓度二次项对发芽红小豆GABA含量有显著影响,且等高线图呈椭圆状。由图7可知,A、D因素在较高水平条件下,响应值随着每个因素水平的增大而增大,当响应值增大到极值后,又逐渐减小。由于GABA是红小豆在发芽时期合成的抗逆环境活性物质,尤其是在胚部分含量较多,随着胚芽发育其不断合成,当胚芽发育成熟其GABA含量便不会明显增加。还有一种观点也认为NaCl浓度增加会促进红小豆合成GABA,尹永祺等[43]研究发现NaCl胁迫下多胺降解途径对大豆发芽富集GABA贡献率大于30%。但盐离子浓度过高,超过植物承受的极限,就会导致GABA的含量降低,最终表现为GABA含量增加到极点后又逐渐减少。由图8可知,NaCl浓度和CaCl2浓度都增加对发芽红小豆GABA含量明显提高,呈相互促进作用。任珺等[44]研究发现CaCl2有助于苦豆子的萌发和生长,有减轻盐胁迫的作用。当外源Na+将细胞质膜上的Ca2+取代,引起Na+浓度增加,Na+/Ca2+比值的平衡遭到破坏,此时外源Ca2+会通过细胞质膜和内膜的钙通道进入细胞质中,促进Ca M区域与钙离子结合提高GAD的活性,从而加快谷氨酸合成GABA[45]。外源Ca2+的施加也能促进细胞对K+和NO3−的吸收,补充细胞内Ca2+的不足,保护膜的稳定性,降低盐胁迫下离子毒害作用[46]。
2.7.3 最优条件的确定及验证
利用Design-Expert 13软件对回归模型进行典型分析得到:发芽时间A=0.810551、CaCl2浓度B=0.179352、L-Glu浓度C=0.21802、NaCl浓度D=0.638318,将四个因素编码值转换成实际值为:发芽时间57.72 h、CaCl2浓度4.36 mmol/L、L-Glu浓度3.22 mg/mL、NaCl浓度65.95 mmol/L,上述为L-Glu联合盐胁迫发芽富集CAPP红小豆GABA的最优工艺参数,在此条件下发芽CAPP红小豆的GABA含量是166.111 mg/100 g,为了验证回归分析的可靠性,将前期回归模型优化工艺参数调整为:发芽时间58 h、CaCl2浓度4.4 mmol/L、L-Glu浓度3.2 mg/mL、NaCl浓度66 mmol/L。按照上述最优发芽条件参数进行三次验证实验,结果得出发芽CAPP红小豆GABA含量的实际值是(160.23±2.91 mg/100 g),理论值与实际值的相对误差仅为3.54%,说明上述模型拟合较好,具有实际应用价值。
3. 结论
本实验使用大气压冷等离子体装置处理红小豆种子联合盐胁迫的方法探究对其发芽富集GABA的影响,发现大气压冷等离子体90 kV,120 Hz,20 min处理的红小豆种子最有利于发芽富集GABA。NaCl、CaCl2和L-Glu都具有诱导豆类发芽富集GABA的作用,且NaCl˃CaCl2˃L-Glu。再利用响应面优化最佳条件为发芽时间58 h、CaCl2浓度4.4 mmol/L、L-Glu浓度3.2 mg/mL、NaCl浓度66 mmol/L,此条件下发芽CAPP红小豆GABA含量为160.23±2.91 mg/100 g,是原料红小豆GABA(22.5±0.2 mg/100 g)的7.12倍。说明CAPP红小豆在发芽过程中,辅以L-谷氨酸和盐处理,可有效提高GABA富集量,进一步证明了萌芽有利于提升红小豆中营养活性成分,为合理利用杂粮豆类资源以及开发高GABA功能食品提供新的思路和理论支持。但对于L-谷氨酸联合盐胁迫CAPP红小豆发芽富集GABA过程中体内的离子浓度及酶活力的变化未进一步探究,为深入了解红小豆发芽富集GABA机制,后续可对GABA合成相关酶的酶活及代谢途径进行研究。
-
![]()
![]()
图 2 CAPP电压对红小豆发芽过程中GABA含量的影响
Figure 2. Effect of CAPP voltage on GABA content during germination of adzuki bean
![]()
图 3 CAPP频率对红小豆发芽过程中GABA含量的影响
Figure 3. Effect of CAPP frequency on GABA content during germination of adzuki bean
![]()
图 4 NaCl对红小豆发芽过程中GABA含量的影响
Figure 4. Effect of NaCl on GABA content during germination of adzuki bean
![]()
图 5 CaCl2对红小豆发芽过程中GABA含量的影响
Figure 5. Effect of CaCl2 on GABA content during germination of adzuki bean
![]()
图 6 L-Glu对红小豆发芽过程中GABA含量的影响
Figure 6. Effect of L-Glu on GABA content during germination of adzuki bean
![]()
图 7 发芽时间和NaCl浓度交互作用的响应面图
Figure 7. Response surface plots of the interaction between germination time and NaCl concentration
![]()
图 8 CaCl2和NaCl浓度交互作用的响应面图
Figure 8. Response surface plots of the interaction between CaCl2 and NaCl concentrations
表 1 Box-Behnken试验因素及水平
Table 1 Factors and levels of Box-Behnken experiment
水平 因素 A 发芽时间
(h)B CaCl2浓度
(mmol/L)C L-Glu浓度
(mg/mL)D NaCl浓度
(mmol/L)−1 36 2 1 25 0 48 4 2 50 1 60 6 3 75  下载: 导出CSV
下载: 导出CSV
表 2 响应面试验设计及结果
Table 2 Response surface design and results
实验号 因素 GABA含量(mg/100 g) A B C D 1 1 1 0 0 146.92±5.08 2 1 0 0 -1 137.74±6.31 3 0 0 0 0 157.01±7.06 4 −1 0 0 1 114.76±1.09 5 0 −1 −1 0 130.07±2.78 6 0 0 0 0 160.34±2.09 7 0 0 0 0 159.32±4.88 8 0 0 0 0 152.48±3.38 9 −1 1 0 0 128.94±4.49 10 −1 0 −1 0 123.56±4.74 11 0 1 0 1 151.84±6.19 12 −1 −1 0 0 119.56±4.89 13 0 −1 1 0 120.30±3.97 14 1 0 1 0 139.03±5.92 15 0 0 1 −1 122.17±6.28 16 0 1 0 −1 125.91±4.86 17 1 0 0 1 166.11±6.38 18 0 0 −1 −1 133.12±6.96 19 0 −1 0 1 143.23±5.95 20 1 −1 0 0 141.22±3.39 21 0 −1 0 −1 136.52±4.18 22 −1 0 1 0 121.96±7.12 23 0 0 0 0 155.92±5.02 24 0 1 −1 0 142.95±5.25 25 −1 0 0 −1 122.77±3.29 26 1 0 −1 0 154.75±4.69 27 0 0 −1 1 136.78±2.94 28 0 1 1 0 137.11±5.86 29 0 0 1 1 157.37±7.75
下载: 导出CSV
表 3 响应面回归模型方差分析
Table 3 Analysis of variance with response surface regression model
来源 偏平方和 自由度 均方和 F值 P值 显著性 模型 5583.97 14 398.85 12.30 <0.0001 significant A 1981.87 1 1981.87 61.11 <0.0001 B 152.44 1 152.44 4.70 0.0479 C 45.27 1 45.27 1.40 0.2571 D 703.19 1 703.19 21.68 0.0004 AB 3.38 1 3.38 0.1043 0.7516 AC 49.86 1 49.86 1.54 0.2354 AD 330.96 1 330.96 10.21 0.0065 BC 3.87 1 3.87 0.1194 0.7348 BD 92.34 1 92.34 2.85 0.1137 CD 248.79 1 248.79 7.67 0.0150 A² 926.77 1 926.77 28.58 0.0001 B² 791.57 1 791.57 24.41 0.0002 C² 891.33 1 891.33 27.49 0.0001 D² 423.43 1 423.43 13.06 0.0028 残差 454.01 14 32.43 失拟项 415.86 10 41.59 4.36 0.0843 not significant 纯误差 38.15 4 9.54 总和 6037.97 28
下载: 导出CSV
-
[1] 林杨, 唐琦勇, 楚敏, 等. γ-氨基丁酸的功能、生产及食品应用研究进展[J]. 中国调味品,2021,46(6):173−179 LIN Y, TANG Q Y, CHU M, et al. Research progress on function, production and food application of γ-aminobutyric acid[J]. China Condiment,2021,46(6):173−179.
[2] IKRAM A, SAEED F, AFZAAL M, et al. Nutritional and end-use perspectives of sprouted grains: A comprehensive review[J]. Food Science & Nutrition,2021,9(8):4617−4628.
[3] 周一峰, 陈治然. 不同发芽条件对发芽花豆γ-氨基丁酸含量的影响[C]//杭州: 中国食品科学技术学会第十一届年会论文摘要集,2014:247−248 ZHOU Y F, CHEN Z R. Effect of different germination conditions on the γ-aminobutyric acid content of germinating flowering beans[C]// Hangzhou: Abstracts of the 11th Annual Conference of the Chinese Society of Food Science and Technology,2014:247−248.
[4] 左娜, 陈洁, 吕莹果, 等. 豆类发芽富集GABA的研究[J]. 粮食与油脂,2016,29(6):29−32 ZUO N, CHEN J, LÜ Y G, et al. Research on enrichment of GABA by bean sprouting[J]. Cereals & Oils,2016,29(6):29−32.
[5] JIANG X J, XU Q P, ZHANG A W et al. Optimization of γ-aminobutyric acid (GABA) accumulation in germinating adzuki beans ( Vigna angularis) by vacuum treatment and monosodium glutamate, and the molecular mechanisms[J]. Frontiers in Nutrition,2021,8:693862. doi: 10.3389/fnut.2021.693862
[6] 吕秋洁, 郑经绍, 余宏达, 等. 富含GABA和花色苷的发芽紫糙米加工工艺研究[J]. 热带作物学报,2021,42(1):220−229 LÜ Q J, ZHENG J S, YU H D, et al. Processing technology of germinating purple brown rice rich in γ-aminobutyric acid (GABA) and anthocyanin[J]. Chinese Journal of Tropical Crops,2021,42(1):220−229.
[7] 曾晴, 谢菲, 尹京苑, 等. 大豆发芽富集 γ-氨基丁酸的培养液组分优化及盐胁迫下的富集机理[J]. 食品科学,2017,38(12):96−103 ZENG Q, XIE F, YIN J Y, et al. Optimization of medium composition for γ-aminobutyric acid accumulation in germinated soybean and mechanism of γ-aminobutyric acid accumulation under salt stress[J]. Food Science,2017,38(12):96−103.
[8] ZHANG L C, HAO N, LI W J, et al. Effect of ultrasonic induction on the main physiological and biochemical indicators and γ-aminobutyric acid content of maize during germination[J]. Foods,2022,11(9):1358. doi: 10.3390/foods11091358
[9] YANG R Q, HUI Q R, FENG X Y, et al. The mechanism of freeze-thawing induced accumulation of γ-aminobutyric acid in germinated soybean[J]. Journal of the Science of Food and Agriculture,2020,3:1099−1105.
[10] RENÁTA Š, ĽUDMILA S, ĽUDMILA H, et al. Evaluation of the impact of cold atmospheric pressure plasma on soybean seed germination[J]. Plants,2021,10:177. doi: 10.3390/plants10010177
[11] 李艾霖. 发芽甜荞中GABA富集技术的研究[D]. 晋中:山西农业大学, 2018 LI A L. Study on technology of GABA accumulation in germinate buckwheat[D]. Jinzhong:Shanxi Agricultural University, 2018.
[12] DUMONT H J, ADRIAENS E. Atmospheric cold plasma as new strategy for foods processing-an overview[J]. Innovative Romanian Food Biotechnology,2014,96(1):88−90.
[13] PRIATAMA R A, PERVITASARI A N, PARK S, et al.Current advancements in the molecular mechanism of plasma treatment for seed germination and plant growth[J]. International Journal of Molecular Sciences,2022,23(9):4609. doi: 10.3390/ijms23094609
[14] LIU H K, ZHANG X W, CUI Z Y, et al. Cold plasma effects on the nutrients and microbiological quality of sprouts[J]. Food Research International,2022,159:111655. doi: 10.1016/j.foodres.2022.111655
[15] BOURKE P, ZIUZINA D, BOEHM D, et al. The potential of cold plasma for safe and sustainable food production[J]. Trends in Biotechnology,2018,36(6):615−626. doi: 10.1016/j.tibtech.2017.11.001
[16] CHEN H H, CHANG H C, CHEN Y K, et al. An improved process for high nutrition of germinated brown rice production: Low-pressure plasma[J]. Food Chemistry,2016,191:120−127. doi: 10.1016/j.foodchem.2015.01.083
[17] ŠIMONČICOVÁ J, KRYŠTOFOVÁ S, MEDVECKÁ V, et al. Technical applications of plasma treatments: Current state and perspectives[J]. Applied Microbiology and Biotechnology,2019,13(103):5117−5129.
[18] 姜秀杰, 张桂芳, 张东杰. 真空协同发芽富集豌豆 γ-氨基丁酸的工艺优化研究[J]. 食品科技,2020,45(5):58−63 JIANG X J, ZHANG G F, ZHANG D J. Optimization of vacuum and germination treatment on γ-aminobutyric acid accumulation in pea[J]. Food Science and Technology,2020,45(5):58−63.
[19] 苏欢. 利用选择性基因分型技术和残余杂合系群体定位大豆生育期和分枝数相关QTL[D]. 聊城:聊城大学, 2017 SU H. Using selective genotyping and residual heterozygous line to identify soybean growth period and branch number OTL[D]. Liaocheng:Liaocheng University, 2017.
[20] BOUCHÉ N, FROMM H. GABA in plants: Just a metabolite?[J]. Trends in Plant Science,2004,9(3):110−115. doi: 10.1016/j.tplants.2004.01.006
[21] 何林阳, 杨杨, 陈凤莲, 等. 响应面法优化发芽糙米生物活性物质提取工艺[J]. 食品安全质量检测学报,2022,13(1):199−207 doi: 10.3969/j.issn.2095-0381.2022.1.spaqzljcjs202201027 HE L Y, YANG Y, CHEN F L, et al. Optimization of extraction process of bioactive substances from germinated brown rice by response surface methodology[J]. Journal of Food Safety & Quality,2022,13(1):199−207. doi: 10.3969/j.issn.2095-0381.2022.1.spaqzljcjs202201027
[22] FILATOVA I, AZHARONOK V, KADYROV M. Rf and microwave plasma application for pre-sowing caryopsis treatments[J]. Publication Astron Obs. Belgrade,2010,89:289−292.
[23] MCDONALD M B. Seed germination and seedling establishment[M]. Madison:American Society of Agronomy, 1994:37−60.
[24] BORMASHENKO E, GRYNYOV R, BORMASHENKO Y, et al. Cold radiofrequency plasma treatment modifies wettability and germination speed of plant seeds[J]. Scientific Reports,2012(2):741.
[25] CHEN H H, CHEN Y K, CHANG H C. Evaluation of physicochemical properties of plasma treated brown rice[J]. Food Chemistry,2012,135(1):74−79. doi: 10.1016/j.foodchem.2012.04.092
[26] ALVES JUNIOR C, DE OLIVEIRA VITORIANO J, DA SILVA DL, et al. Water uptake mechanism and germination of Erythrina velutina seeds treated with atmospheric plasma[J]. Scientific Reports,2016,6:33722. doi: 10.1038/srep33722
[27] 王佳媚, 黄明明, 乔维维, 等. 冷源等离子体冷杀菌技术及其在食品中的应用研究[J]. 中国农业科技导报,2015,17(5):55−62 WANG J M, HUANG M M, QIAO W W, et al. Disinfection technology of cold plasma and its application in food[J]. Journal of Agricultural Science and Technology,2015,17(5):55−62.
[28] 何瑞, 童家赟, 张晓丽, 等. 大气压等离子体处理对穿心莲种子萌发及幼苗的影响初报[J]. 广东农业科学,2011,38(16):23−25 doi: 10.3969/j.issn.1004-874X.2011.16.009 HE R, TONG J B, ZHANG X L, et al. Preliminary study of effect of atmospheric pressure plasma on the germination and seedling of Andrographis paniculate[J]. Guangdong Agricultural Sciences,2011,38(16):23−25. doi: 10.3969/j.issn.1004-874X.2011.16.009
[29] BUTSCHER D, VAN LOON H, WASKOW A, et al. Plasma inactivation of microorganisms on sprout seeds in a dielectric barrier discharge[J]. International Journal of Food Microbiology,2016,238:222−232. doi: 10.1016/j.ijfoodmicro.2016.09.006
[30] VOLKOV A G, BOOKAL A, HAIRSTON J S, et al. Radio frequency plasma capacitor can increase rates of seeds imbibition, germination, and radicle growth[J]. Functional Plant Biology,2021,48(3):312−320. doi: 10.1071/FP20293
[31] AL-QURAAN N A, SARTAWE F A, QARYOUTI M M. Characterization of γ-aminobutyric acid metabolism and oxidative damage in wheat ( Triticum aestivum L.) seedlings under salt and osmotic stress[J]. Journal of Plant Physiology,2013,170(11):1003−1009. doi: 10.1016/j.jplph.2013.02.010
[32] XING S G, JUN Y B, HAU Z W, et al. Higher accumulation of gamma-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots[J]. Plant Physiology and Biochemistry,2007,45(8):560−566. doi: 10.1016/j.plaphy.2007.05.007
[33] YANG R Q, WANG M, FENG X Y, et al. AMADH inhibitor optimization and its effects on GABA accumulation in soybean sprouts under NaCl-CaCl2 treatment[J]. 3 Biotech,2019,9(5):184. doi: 10.1007/s13205-019-1715-7
[34] ZHAO Y Y, XIE C, WANG P, et al. GABA regulates phenolics accumulation in soybean sprouts under NaCl stress[J]. Antioxidants (Basel),2021,10(6):990. doi: 10.3390/antiox10060990
[35] FALCINELLI B, SILEONI V, MARCONI O, et al. Germination under moderate salinity increases phenolic content and antioxidant activity in rapeseed ( Brassica napus var oleifera Del.) sprouts[J]. Molecules,2017,22(8):1377. doi: 10.3390/molecules22081377
[36] BAUM G, CHEN Y, ARAZI T, et al. A plant glutamate decarboxylase containing a calmodulin binding domain. Cloning, sequence, and functional analysis[J]. Journal of Biological Chemistry,1993,26(268):19610−19617.
[37] WANG Y S, LUO Z S, MAO L C, et al. Contribution of polyamines metabolism and GABA shunt to chilling tolerance induced by nitric oxide in cold-stored banana fruit[J]. Food Chemistry,2016,197:333−339. doi: 10.1016/j.foodchem.2015.10.118
[38] 王姗姗, 刘小娇, 胡赟, 等. 植物中 γ-氨基丁酸的代谢及富集机制[J]. 安徽农业科学,2020,48(24):9−12 doi: 10.3969/j.issn.0517-6611.2020.24.003 WANG S S, LIU X J, HU B, et al. Metabolism and enrichment mechanism of γ-aminobutyric acid in plants[J]. Journal of Anhui Agricultural Sciences,2020,48(24):9−12. doi: 10.3969/j.issn.0517-6611.2020.24.003
[39] 朱云辉, 郭元新. 响应面法优化发芽苦荞富集 γ-氨基丁酸的培养条件[J]. 西北农林科技大学学报(自然科学版),2016,44(11):141−148 ZHU Y H, GUO Y X. Optimizing culture conditions for GABA accumulation in germinated buckwheat based on response surface methodology[J]. Journal of Northwest A & F University (Natural Science Edition),2016,44(11):141−148.
[40] 王斌, 丁俊胄, 贾才华, 等. 环境胁迫植物富集 γ-氨基丁酸的研究进展[J]. 食品工业科技,2018,39(18):342−346,352 WANG B, DING J Z, JIA C H, et al. Research progress on enrichment of γ-aminobutyric acid in plants under environmental stress[J]. Science and Technology of Food Industry,2018,39(18):342−346,352.
[41] 顾振新, 蒋振晖. 食品原料中 γ-氨基丁酸(GABA)形成机理及富集技术[J]. 食品与发酵工业,2002(10):65−69 doi: 10.3321/j.issn:0253-990X.2002.10.016 GU Z X, JIANG Z H. Mechanism of γ-aminobutyric acid (GABA) formation and enrichment technology in food materials[J]. Food and Fermentation Industries,2002(10):65−69. doi: 10.3321/j.issn:0253-990X.2002.10.016
[42] 陈惠, 杨润强, 韩永斌, 等. 发芽蚕豆富集 γ-氨基丁酸的培养液组分优化[J]. 中国粮油学报,2011,26(11):27−31 CHEN H, YANG R Q, HAN Y B, et al. Optimization of culture medium components for the enrichment of γ-aminobutyric acid in germinating broad beans[J]. Journal of the Chinese Cereals and Oils Association,2011,26(11):27−31.
[43] 尹永祺. NaCl及其联合Ca2+处理下发芽大豆生理变化与GABA富集调控机理[D]. 南京:南京农业大学, 2014 YIN Y Q. Study on the physiological change and the regulation mechanism of GABA accumulation in germinating soybean under NaCl and NaCl-Ca2+ treatment[D]. Nanjing: Nanjing Agricultural University, 2014.
[44] 任珺, 孙梦洁, 张照桤, 等. 外源钙对盐胁迫下苦豆子( Sophora alopecuroides)种子萌发和幼苗生长的影响[J]. 中国沙漠,2019,39(1):105−109 REN J, SUN M J, ZHANG Z Q, et al. Seed germination and seedling growth of Sophora alopecuroides under saline alkali stress[J]. Journal of Desert Research,2019,39(1):105−109.
[45] 尹永祺, 李童, 王淑雯, 等. 钙离子通道抑制剂处理下发芽大豆主要生理生化和 γ-氨基丁酸的代谢变化[J]. 食品工业科技,2016,37(16):122−126 YIN Y Q, LI T, WANG S W, et al. Physiological change and the regulation of γ-aminobutyric acid accumulation in germinated soybean under inhibitors of calcium channel treatment[J]. Science and Technology of Food Industry,2016,37(16):122−126.
[46] 郑秀芳, 张超强. 外源钙对盐胁迫下马铃薯试管苗生长和相关生理特性的影响[J]. 西北农业学报,2015,24(6):97−102 doi: 10.7606/j.issn.1004-1389.2015.06.015 ZHENG X F, ZHANG C Q. Effects of exogenous Ca(NO3)2 on growth and physiological characteristics of potato test-tube seedlings under salt stress[J]. Acta Agriculturae Boreali-occidentalis Sinica,2015,24(6):97−102. doi: 10.7606/j.issn.1004-1389.2015.06.015
 下载:
下载:










计量
- 文章访问数: 85
- HTML全文浏览量: 30
- PDF下载量: 19
