


Effect of β-Nicotinamide Mononucleotide on the Growth Promotion of Caenorhabditis elegans and Its Mechanism
-
摘要: 目的:研究β-烟酰胺单核苷酸(β-nicotinamide mononucleotide,NMN)对秀丽隐杆线虫生长的影响与作用机制。方法:将线虫随机分为空白对照组和不同质量浓度NMN实验组(0.1、0.5、1、5、10 mg/mL),通过测定线虫在成年后13 d内的体长变化以评估NMN对线虫生长的影响;基于转录组学探索NMN(1 mg/mL)促线虫生长的作用机制;并采用实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)验证测序结果。结果:经0.1、0.5、1、5、10 mg/mL NMN喂食线虫后,相较空白对照组,线虫的平均体长在4~13 d均得到显著增加(P<0.05)。转录组测序分析筛选出263个差异表达基因(differentially expressed genes,DEGs),基因本体论(gene ontology,GO)分析发现这些DEGs主要涉及胶原蛋白参与的角质层发育和基于角质素的角质层蜕皮周期、基于胶原蛋白和角质素的角质层发育、胶原蛋白三聚体等与生长发育有关的生物学功能。对富集的关键GO中的DEGs构建蛋白互作网络图,并对筛选出的前10个关键核心基因进行qRT-PCR验证,结果基本与转录组一致。结论:NMN可能通过调节胶原蛋白和角质层相关基因的表达促进线虫生长。本研究为功能性食品、保健品等的开发提供了新的思路。Abstract: Objective: To study the effect of β-nicotinamide mononucleotide (NMN) on the growth of Caenorhabditis elegans (C. elegans) and its mechanism. Methods: Nematodes were randomly divided into blank control group and experimental groups with different concentrations of NMN (0.1, 0.5, 1, 5, 10 mg/mL). The impact of NMN on the growth of C. elegans was evaluated by measuring the body length changes of C. elegans in 13 days after adulthood. In addition, transcriptomics was used to explore the mechanism of NMN (1 mg/mL) in promoting nematode growth. Moreover, quantitative real-time polymerase chain reaction (qRT-PCR) was performed to verify the sequencing results. Results: Compared with the blank control group, the average body length of C. elegans was significantly increased after feeding 0.1, 0.5, 1, 5 and 10 mg/mL NMN at 4 to 13 days (P<0.05). Totally 263 differentially expressed genes (DEGs) were identified by transcriptome sequencing analysis. Gene ontology (GO) analysis showed that these DEGs were mainly involved in cuticle development involving collagen, cuticulin-based cuticle molting cycle, collagen and cuticulin-based cuticle development, collagen trimer and other biological functions related to growth and development. The protein interaction network diagram was constructed for the DEGs in the enriched key GOs, then the top 10 key genes were screened out for qRT-PCR verification, and the results were basically consistent with the transcriptome. Conclusion: NMN may promote the growth of C. elegans by regulating the expression of collagen and cuticle related genes. This study provides new ideas for the development of functional foods and health products.
-
烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD+)是氧化还原反应的重要辅酶,是能量代谢的核心,NAD+对于所有生命形式的代谢反应至关重要,同时其在调节细胞过程和功能等方面也发挥着多种作用[1-2]。随着年龄的增长,NAD+的水平不断下降,而补充NAD+的前体或者限制NAD+的消耗可以缓解由于NAD+水平降低所引起的机体功能衰退[3-5]。β-烟酰胺单核苷酸(β-nicotinamide mononucleotide,NMN)是一种NAD+的关键前体物质[6],口服NMN能够提升组织NAD+水平,逆转因NAD+不足而造成的线粒体功能异常、活性氧产生、DNA受损以及细胞存活等方面的缺陷[7]。近年来NMN也因其在抗衰老方面的重要应用价值而受到广泛的关注,有研究报道NMN在调节衰老小鼠的肠道结构和缓解功能衰退方面具有一定潜力[8];Kim等[9]通过人体临床试验发现午后服用NMN,持续干预12周,老年人受试者的疲劳程度显著减轻,下肢身体技能得到明显提升;另外NMN还可以作为抗黑色素生成剂,有效减少老化的重组人体皮肤细胞中黑色素的产生[10];此外,NMN在抗病毒感染[11]、抗炎[12]、抗过敏[13]、降血糖[14]等方面也表现出一定的效果。越来越多的研究证明NMN在改善机体功能方面起着重要作用,然而目前关于NMN对机体生长的影响鲜有报道。
生长发育是机体重要的生命活动,发育迟缓易诱发精神类疾病和一系列行为问题[15-16],而生长又是发育的基础,研究发现通过营养干预可以有效减少生长迟缓[17]。秀丽隐杆线虫(Caenorhabditis elegans,C. elegans)是一种模式生物,与人类基因的同源性达60%~80%[18],因其具有个体结构简单、生活史短、遗传背景清楚、易于观察等特点,常作为动物模型广泛应用于发育生物学、食品安全研究中[19-20],如程雷[21]以C. elegans作为研究对象,探讨了稳态磁场对C. elegans早期胚胎细胞及个体发育的影响;Pradhan等[22]利用C. elegans评估了增塑剂邻苯二甲酸二(2-乙基己基)酯的毒性及作用机理;石钰等[23]研究了食品包装禁用材料双酚A对C.elegans生长发育及生殖毒性的影响。
转录组测序是近几年发展起来的一种高通量测序技术,通过基因表达水平的变化及调控规律为作用机制的解析提供有效手段[24]。其在揭示中药、食品提取物等作用机理方面有诸多应用,如叶丽云等[24]通过转录组学分析了赤芝多糖有效预防急性酒精肝损伤可能的作用机制;何秋玲等[25]通过转录组学挖掘了异麦芽酮糖减少小鼠肝脏脂肪堆积的关键基因;张婷婷等[26]利用转录组学阐释了普洱茶茶褐素对代谢综合征大鼠肝脏基因表达谱的影响。
基于此,本研究以C. elegans作为动物模型,探究了NMN对线虫生长的影响,并通过转录组测序探讨NMN促线虫生长的作用机制,以期为拓展NMN的应用、开发促生长功能性食品、保健品、药品或饲料等提供新的理论依据。
1. 材料与方法
1.1 材料与仪器
秀丽隐杆线虫(Caenorhabditis elegans,C. elegans)野生型N2、大肠杆菌(Escherichia coli,E. coli)尿嘧啶缺陷型OP50 由四川大学生命科学学院资源微生物学及微生物生物技术四川省重点实验室提供。
β-烟酰胺单核苷酸 上海麦克林生化科技股份有限公司;胆固醇 北京索莱宝科技有限公司;LB肉汤、LB琼脂 青岛高科技工业园海博生物技术有限公司;5-氟-2′-脱氧尿嘧啶核苷(floxuridine, FUDR) 美国Sigma公司;逆转录试剂、TB Green 宝生物工程(大连)有限公司;其余生化试剂(分析纯) 成都金山化学试剂有限公司。
BSA124S电子天平 德国Sartorius公司;AFZ-1002-U超纯水系统 美国Aquapro公司;LDZX-50KBS立式压力蒸汽灭菌锅 上海申安医疗器械厂;1389生物安全柜 美国Thermo公司;DHP-9052电热恒温培养箱 上海一恒科学仪器有限公司;SKY-211C恒温摇床 上海苏坤实业有限公司;5804R高速台式离心机 德国Eppendorf公司;SMZ-B2体式显微镜 重庆奥特光学仪器有限公司;DP73光学显微镜 日本Olympus公司;NovaSeq6000测序仪 美国Illumina公司;QuantStudio 3实时荧光定量PCR仪 美国Thermo公司。
1.2 实验方法
1.2.1 待测样品的制备
称取适量NMN粉末,用无菌水充分溶解后配制成质量浓度为10 mg/mL的NMN母液,经0.22 μm微孔滤膜过滤除菌后,按照倍比稀释法依次配制终浓度为5、1、0.5、0.1 mg/mL的NMN溶液,4 ℃保存备用。
1.2.2 主要培养基的配制
线虫生长培养基(nematode growth medium,NGM)的配制[27]:称取3.0 g NaCl,8.5 g琼脂粉,1.25 g胰蛋白胨于500 mL蓝口瓶中,加入488 mL超纯水充分混匀,121 ℃灭菌20 min,待冷却至55 ℃左右,再分别加入0.5 mL 5 mg/mL胆固醇乙醇溶液、12 mL 1 mol/L磷酸盐缓冲液、0.5 mL 1 mol/L CaCl2溶液、0.5 mL 1 mol/L MgSO4溶液,混匀后迅速倒入6 cm或9 cm无菌平板中,待培养基凝固后备用。
LB培养基的配制:LB固体及液体培养基的配制方法参照试剂说明书。
NGM-OP50培养板的制备(用于线虫常规培养)[27]:将活化好的E. coli OP50接种至LB液体培养基中,37 ℃摇床培养16 h左右,吸取130 μL E. coli OP50菌悬液均匀涂布于9 cm NGM平板上,37 ℃过夜培养,备用。
含待测样品NGM-OP50培养板的制备(用于线虫体长的测定及转录组样本收集):取100 μL经灭活处理的E. coli OP50菌悬液均匀涂布于6 cm NGM平板上,室温放置过夜,次日分别吸取100 μL无菌水(空白对照组)、不同质量浓度NMN溶液(实验组)再次涂布于含灭活E. coli OP50的6 cm NGM平板上,自然干燥后备用。
M9缓冲液的配制[27]:称取1.5 g KH2PO4,3 g Na2HPO4,2.5 g NaCl于500 mL蓝口瓶中,加入500 mL超纯水充分混匀,121 ℃灭菌20 min,冷却后加入1 mL 1 mol/L MgSO4溶液,备用。
1.2.3 线虫常规培养
取−80 ℃冻存的线虫于室温下自然解冻,用移液枪轻轻吹吸混匀后接种至9 cm NGM-OP50培养板,20 ℃恒温培养箱中培养数天后可对线虫进行转接扩繁,待线虫发育成怀卵线虫后可对线虫进行同步化处理[27]。
1.2.4 线虫同步化
用3 mL M9缓冲液冲洗NGM-OP50培养板上已发育至成虫的怀卵线虫,并将其转移至15 mL离心管中,按顺序依次加入0.8 mL NaClO溶液、0.4 mL 5 mg/mL NaOH溶液,来回颠倒混匀数次后,置于离心机中5500 r/min 离心1 min,弃上清,在无菌条件下加入4 mL M9缓冲液,混匀后5500 r/min再次离心1 min,弃上清,随后加入4 mL M9缓冲液,吹吸混匀后将含虫卵的液体转移至3.5 cm培养皿中,20 ℃恒温培养箱中过夜孵化[27]。
1.2.5 线虫处理及体长的测定
将过夜孵化的L1期线虫转接至含无菌水或不同质量浓度待测样品的NGM-OP50培养板中,待线虫长至L4期,按照每个6 cm NGM-OP50培养板中30条线虫的比例进行随机分配转移。首先,将L4期线虫转移至同时含有终浓度为40 µmol/L FUDR的NGM-OP50培养板中培养2 d,以防止线虫生殖;随后,将线虫转移至不含FUDR的NGM-OP50培养板中继续培养,线虫成年后1~5 d内需每天将其转移至新鲜的NGM-OP50培养板中,线虫成年5 d后每2 d将其转移至新鲜的NGM-OP50培养板中[28-29]。设置无菌水空白对照组(0 mg/mL)和不同质量浓度NMN实验组(0.1、0.5、1、5、10 mg/mL),分别于线虫成年的第4、7、10、13 d采用显微镜进行拍照并测量线虫虫体的长度。
1.2.6 线虫组学样本收集
根据线虫体长的测定结果,实验组加入促线虫生长效果最好质量浓度的NMN溶液,对照组加入等量的无菌水,每组设置3个平行,分别于无菌水或待测样品处理线虫的第0 d(A00组)、第7 d(对照组:A07组、实验组:AN7组)和第13 d(对照组:A013组、实验组:AN13组),用M9缓冲液将NGM-OP50培养板上的线虫转移至离心管中,清洗3次,前2次采用5500 r/min离心3 min,第3次采用13000 r/min离心10 min,吸净上清,液氮速冻30 min后置于-80 ℃保存备用[30-31]。
1.2.7 转录组测序与分析
将上述各组线虫样本送至上海美吉生物医药科技有限公司进行RNA提取、质检、文库构建及上机测序。采用Trizol法提取线虫的总RNA,NanoDrop2000检测RNA的质量浓度和纯度,琼脂糖凝胶电泳检测RNA的完整性,质检合格的样本采用NovaSeq6000进行转录组测序。将下机后的原始测序数据(Raw data)进行过滤,得到的高质量测序数据(Clean data)通过Hisat2软件[32]与线虫的参考基因组(参考基因组来源:http://metazoa.ensembl.org/Caenorhabditis_elegans/Info/Index)进行比对,获得可以定位到基因组的数据(Mapped data)用于后续基因表达量的计算。采用RSEM软件对基因的表达水平进行定量分析,并通过DESeq2软件[33]按照P<0.05和|log2FC|≥1的标准筛选差异表达基因(Differentially Expressed Genes,DEGs)。再利用Goatools软件对DEGs进行基因本体论(Gene Ontology,GO)分析[34]以获得关键富集的生物学功能,并进一步对显著富集GO条目中的DEGs进行蛋白互作网络分析以获得关键核心基因。
1.2.8 实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)
采用PrimeScript™RT试剂盒将各组RNA样本反转录成cDNA,以act-1作为内参基因,使用QuantStudio 3实时荧光定量PCR仪检测上述关键核心基因的表达,并根据2−ΔΔCt法计算基因的相对表达量。引物序列(表1)参考相关文献[35]或利用Primer Premier 5.0软件进行设计。
表 1 关键核心基因引物序列Table 1. Primer sequences of key genes基因 上游引物(5’-3’) 下游引物(5’-3’) col-107 AGCCATCGTTATTGGAGCGTCTTTC AGGCTTCATTGGCGGTGTTTCTG dpy-13 CACAGAAGCCTTGCGAGGAGATC TCCTGGCTGTCCTGGTTGTCC col-125 GAAACCGTACCGCCCGTCAAG TGGAAGGCAGCAAGCATCACATC dpy-4 AACCGTACCGCCCGTTCTACC GCTCCTGGCTTTCCTGGCTTTC col-180 CCGCACTCAAGTTCTCGCTAAGG CAGCAGCAGAGACACCGCATC col-168 TCTCCTTCACTGCCACTTTG TCCAGCAGCAGATACTCCAC col-167 CGTCTCCTTCACTGCCACA ACTCCACATCCGCAGCAT sqt-1 AGCGTGTCCGTCGTCAATATGAAG CTGGTGGAACTGCTGGTGGTTG col-94 CTTCCCGTTTCGCCCGTCAAG GCATCCAGAGCATGATCCTCCTTG col-144 TGCCAATGCGAGCCAACTAAGC AGGTGAGTGGAGCGAAGGTAGC act-1 TCGGTATGGGACAGAAGGAC CATCCCAGTTGGTGACGATA 1.3 数据处理
采用GraphPad Prism 8.0软件对线虫体长检测的结果进行单因素方差分析,实验结果以平均值±标准差表示,图表中各组间的显著性差异用星号表示,P<0.05表示结果具有统计学意义。
2. 结果与分析
2.1 NMN对线虫生长的影响
体长是衡量线虫个体生长发育最为常见的指标[36-37]。为探讨NMN对线虫生长的影响,采用不同质量浓度的NMN对线虫体长的作用效果进行了评估。由表2可知,采用质量浓度为0.1、0.5、1、5、10 mg/mL的NMN分别喂食线虫,线虫的平均体长在4~13 d均显著高于其相应的空白对照组(P<0.05),表明0.1、0.5、1、5、10 mg/mL的NMN可以促进线虫体长的增加;在相同处理时间条件下,线虫的平均体长随着NMN质量浓度的增加整体呈现先升高后降低的趋势。其中,当采用质量浓度为1 mg/mL的NMN喂食线虫时,线虫的平均体长增加最大(图1),分别在第4、7、10、13 d相较其对应的空白对照组提高了62.82%、54.02%、25.46%、35.24%(表2)。以上数据表明,实验组中所有剂量的NMN均能促进线虫的生长,使其在体长上表现出明显的增加。
表 2 NMN对线虫体长的影响Table 2. Effect of NMN on the body length of C. elegans组别 平均体长(μm) 第4 d 第7 d 第10 d 第13 d 空白组 1097.21±58.30 1358.12±226.87 1744.96±179.89 1724.14±197.68 0.1 mg/mL NMN 1690.08±127.68**** 1874.49±175.45*** 2010.70±115.03* 1920.16±130.65* 0.5 mg/mL NMN 1682.49±73.88**** 1845.79±246.27*** 2121.82±138.26*** 2124.91±149.15**** 1 mg/mL NMN 1786.50±74.23**** 2091.75±62.33**** 2189.21±165.56**** 2331.77±94.70**** 5 mg/mL NMN 1779.99±140.23**** 1994.70±221.00**** 2133.55±252.82**** 2127.99±113.93**** 10 mg/mL NMN 1580.63±118.64**** 2048.12±206.78**** 2176.72±148.28**** 2112.67±165.67**** 注:*与空白组相比,P<0.05;***与空白组相比,P<0.001;****与空白组相比,P<0.0001。 ![]() 图 1 NMN喂食线虫第13 d的照片注:A、B、C、D、E、F分别为空白组、0.1、0.5、1、5、10 mg/mL NMN处理组。Figure 1. Photograph of C. elegans on day 13 of NMN treatment
图 1 NMN喂食线虫第13 d的照片注:A、B、C、D、E、F分别为空白组、0.1、0.5、1、5、10 mg/mL NMN处理组。Figure 1. Photograph of C. elegans on day 13 of NMN treatment2.2 线虫转录组测序数据统计与质量评估
基于NMN对线虫生长的显著促进效果,为了进一步研究NMN对线虫基因表达水平的影响,探讨NMN促线虫生长可能的作用机制,利用转录组测序对1 mg/mL NMN处理第0 d(A00组)、第7 d(对照组:A07组、实验组:AN7组)和第13 d(对照组:A013组、实验组:AN13组)的线虫样本进行了分析。由表3可知,各组线虫样本经转录组测序共获得810840930个Raw reads,质控后共获得798281728个Clean reads,所有样本与线虫参考基因组的比对率均大于95%,且Q20均大于97%,Q30均大于92%。一般认为Q20大于85%,Q30大于80%即可满足测序的质量要求[38]。由此可见,此次测序数据的数量和质量均已达到后续分析的要求。
表 3 线虫样本转录组测序统计Table 3. Transcriptomic sequencing statistics of C. elegans样本编号 Raw reads Clean reads Q20(%) Q30(%) 比对率(%) A001 59049504 57669184 97.31 92.69 95.90 A002 44952038 44653322 97.68 93.36 95.90 A003 53516110 52311272 97.21 92.39 95.97 A071 55185606 53795278 97.47 93.01 96.48 A072 43503612 43215388 97.76 93.57 95.90 A073 56342184 55299230 97.77 93.68 96.49 AN71 54934544 54178138 97.77 93.72 96.46 AN72 57107824 56171120 97.29 92.52 95.98 AN73 61027166 59699564 97.61 93.35 96.20 A0131 44940238 44590662 97.40 92.71 95.56 A0132 55203858 54770834 97.51 92.96 95.78 A0133 64016058 63209158 97.54 93.07 96.08 AN131 44595066 44277946 97.69 93.41 95.61 AN132 59915312 58374528 97.37 92.74 96.30 AN133 56551810 56066104 97.66 93.33 95.85 注:样本编号的最后1位数字1、2、3分别代表该组的3个平行样本。 2.3 差异表达基因的筛选
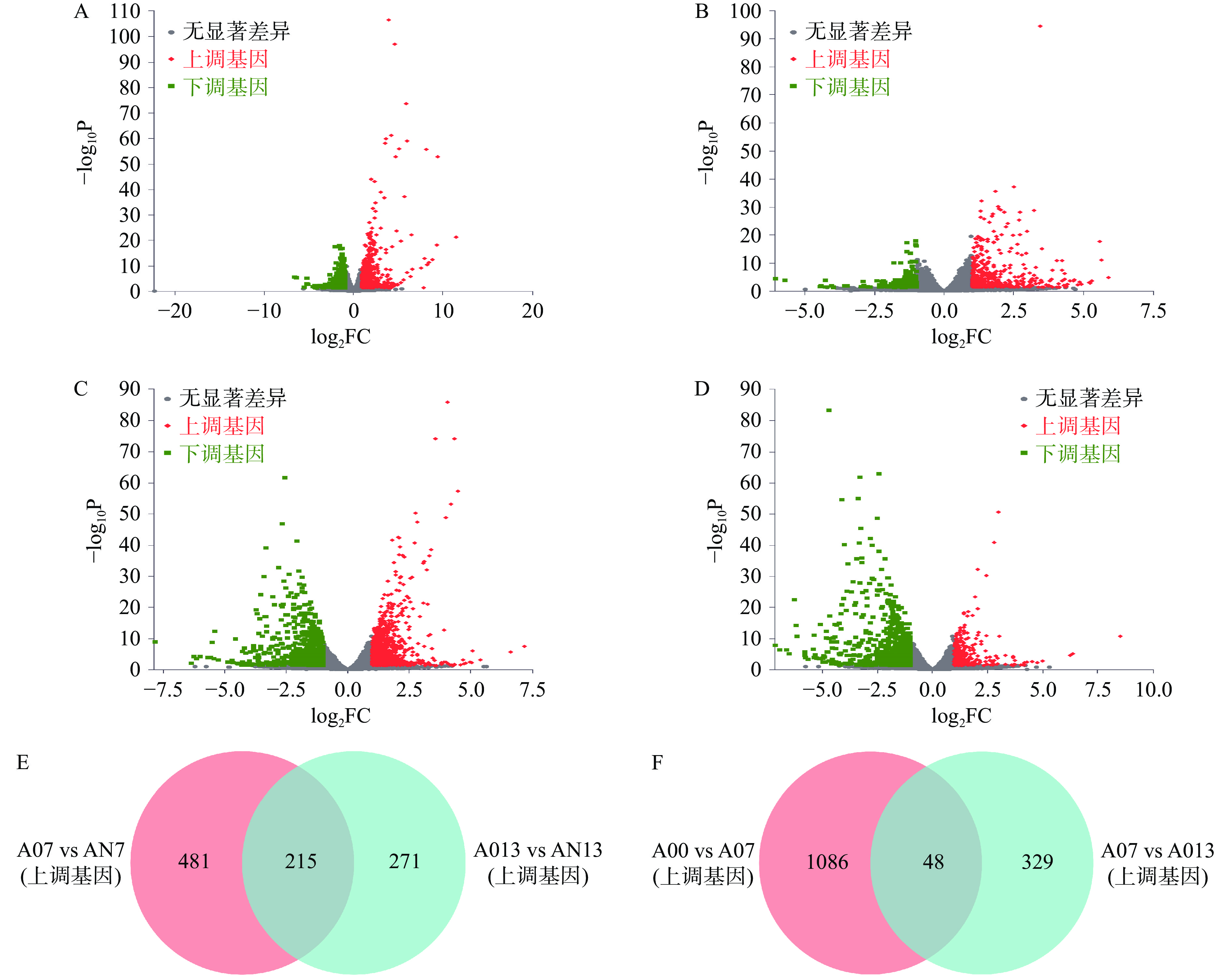
为筛选出DEGs,将A07vsAN7、A013vsAN13、A00vsA07、A07vsA013的转录组数据进行比较分析。由图2A、图2B可知,AN7与A07相比较,共筛选出1334个DEGs,其中上调基因696个,占总DEGs的52.17%,下调基因638个,占总DEGs的47.83%;AN13与A013相比较,共筛选出754个DEGs,其中上调基因486个,占总DEGs的64.46%,下调基因268个,占总DEGs的35.54%。结果表明喂食NMN可以显著改变线虫基因的表达水平,且受NMN影响的线虫上调基因的数目高于下调基因的数目。由图2C、图2D可知,A07与A00相比较,共筛选出1983个DEGs,其中上调基因1134个,下调基因849个;A013与A07相比较,共筛选出1149个DEGs,其中上调基因377个,下调基因772个。结合前期研究发现A07vsAN7和A013vsAN13的共同下调基因主要参与细胞代谢过程,A00vsA07和A07vsA013的共同下调基因主要参与应答反应,并未直接涉及生长发育,综上选择A07vsAN7和A013vsAN13、A00vsA07和A07vsA013的共同上调基因进行后续富集分析,其中A07vsAN7和A013vsAN13的共同上调基因有215个,A00vsA07和A07vsA013的共同上调基因有48个(图2E、图2F)。
![]() 图 2 差异表达基因分析与筛选注:A、B、C、D分别为A07vsAN7、A013vsAN13、A00vsA07、A07vsA013差异表达基因火山图;E为A07vsAN7和A013vsAN13的共同上调差异基因韦恩图;F为A00vsA07和A07vsA013的共同上调差异基因韦恩图。Figure 2. Analysis and screening of differentially expressed genes
图 2 差异表达基因分析与筛选注:A、B、C、D分别为A07vsAN7、A013vsAN13、A00vsA07、A07vsA013差异表达基因火山图;E为A07vsAN7和A013vsAN13的共同上调差异基因韦恩图;F为A00vsA07和A07vsA013的共同上调差异基因韦恩图。Figure 2. Analysis and screening of differentially expressed genes2.4 差异表达基因GO富集分析
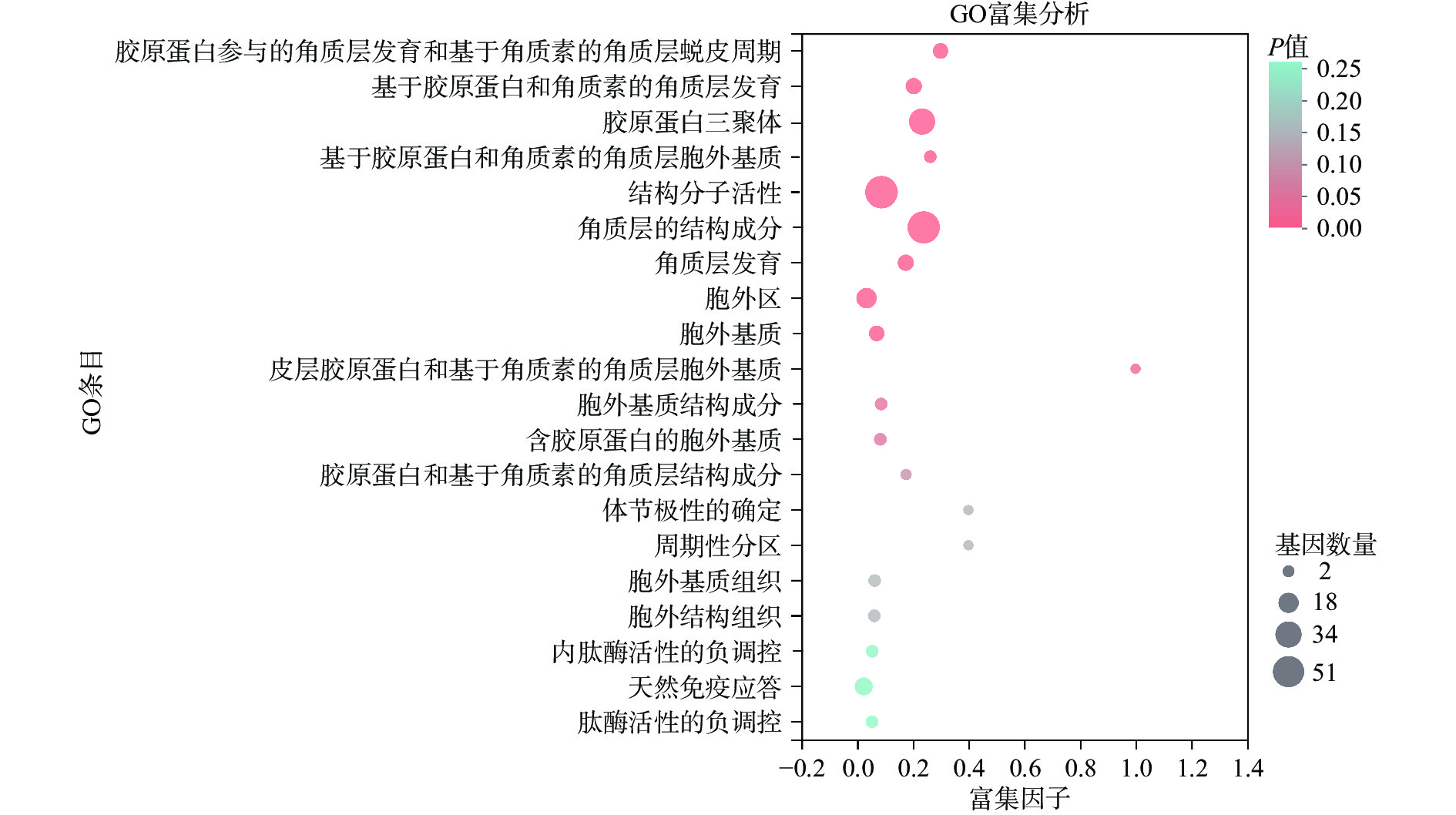
对上述筛选出的263个满足条件的DEGs进行GO富集分析,以获得这些DEGs参与的生物学功能。如图3所示,这些DEGs主要涉及胶原蛋白参与的角质层发育和基于角质素的角质层蜕皮周期、基于胶原蛋白和角质素的角质层发育、胶原蛋白三聚体、基于胶原蛋白和角质素的角质层胞外基质、结构分子活性、角质层的结构成分、角质层发育、胞外区、胞外基质、皮层胶原蛋白和基于角质素的角质层胞外基质、胞外基质结构成分、含胶原蛋白的胞外基质、胶原蛋白和基于角质素的角质层结构成分等,而这些GO条目大多与胶原蛋白和角质层有关,说明NMN喂食线虫可以影响其与胶原蛋白和角质层有关的生物学功能。进一步将关键富集的前10个GO条目绘制GO有向无环图(图4),将GO分成细胞组分、分子功能和生物过程3大功能类别,从图中可以直观地看出各个GO条目之间的从属关系,发现在生物过程类别中,胶原蛋白参与的角质层发育和基于角质素的角质层蜕皮周期、基于胶原蛋白和角质素的角质层发育和角质层发育这3个关键富集的GO条目可以溯源至发育过程这条GO,说明NMN喂食影响了线虫的发育,而NMN可能通过改变这些GO条目中基因的表达促进线虫的生长发育。
![]() 图 3 差异表达基因GO富集分析注:图中圆点的颜色对应不同的P值范围;圆点的大小代表在该GO条目中的基因数量;富集因子表示该GO条目中富集到的基因数量占该GO条目中总基因数量的比值,富集因子越大,表示GO的富集程度越大。Figure 3. GO enrichment analysis of differentially expressed genes
图 3 差异表达基因GO富集分析注:图中圆点的颜色对应不同的P值范围;圆点的大小代表在该GO条目中的基因数量;富集因子表示该GO条目中富集到的基因数量占该GO条目中总基因数量的比值,富集因子越大,表示GO的富集程度越大。Figure 3. GO enrichment analysis of differentially expressed genes![]() 图 4 差异表达基因GO有向无环图Figure 4. GO directed acyclic graph of differentially expressed genes
图 4 差异表达基因GO有向无环图Figure 4. GO directed acyclic graph of differentially expressed genes2.5 差异表达基因蛋白互作网络分析
基于上述关键GO富集分析的结果,利用数据库中基因之间的相互作用关系以构建蛋白互作网络图,从而进一步分析NMN促线虫生长过程中的关键核心基因。如图5所示,图中圆点代表差异表达基因,直线代表不同基因之间具有相互作用关系,不同颜色对应不同的Degree值范围,Degree值越高的基因节点越大,说明与该基因相互作用的基因越多,其在该互作网络中的重要性越强。从图5可以直观地看出,col-107的节点最大,Degree值为25;其次是dpy-13、col-125、dpy-4,Degree值分别为21、21、20;col-180、col-168、col-167、sqt-1、col-94、col-144、col-65、col-77的Degree值均大于15;其中,col-107、dpy-13、col-125、dpy-4、col-168、col-144、col-65是编码线虫表皮相关的胶原蛋白基因[39-41],col-107、col-167、col-77可能与角质层结构构成有关[42-43],dpy-13、sqt-1已被证可以影响线虫的体型[44-45],以上结果提示这些基因可能在NMN促线虫生长方面起着重要的作用。
![]() 图 5 差异表达基因蛋白互作网络图注:图中Degree值代表节点的连通性,节点的颜色对应不同的Degree值范围,Degree值越大,说明与该节点相互作用的基因越多;节点大小代表基因在该网络中的重要程度,节点越大,说明基因在该网络中越重要。Figure 5. Protein-protein interaction network of differentially expressed genes
图 5 差异表达基因蛋白互作网络图注:图中Degree值代表节点的连通性,节点的颜色对应不同的Degree值范围,Degree值越大,说明与该节点相互作用的基因越多;节点大小代表基因在该网络中的重要程度,节点越大,说明基因在该网络中越重要。Figure 5. Protein-protein interaction network of differentially expressed genes2.6 关键核心基因差异表达验证
对蛋白互作网络分析获得的前10个关键核心基因进行qRT-PCR验证,结果显示:当采用质量浓度为1 mg/mL的NMN喂食线虫时,线虫col-107、dpy-13、col-125、dpy-4、col-180、col-168、col-167、sqt-1、col-94、col-144基因mRNA的表达水平在第7 d(图6A)、第13 d(图6B)均明显上调,且差异表达倍数均大于2倍。qRT-PCR验证结果基本与转录组结果一致,表明转录组数据准确可靠。
![]() 图 6 A07vsAN7(A)和A013vsAN13(B)关键核心基因差异表达验证Figure 6. Validation of differential expression of key genes in A07vsAN7 (A) and A013vsAN13 (B)
图 6 A07vsAN7(A)和A013vsAN13(B)关键核心基因差异表达验证Figure 6. Validation of differential expression of key genes in A07vsAN7 (A) and A013vsAN13 (B)3. 讨论与结论
本研究以秀丽隐杆线虫作为动物模型,利用体长测定的方法探究了NMN对线虫生长的影响。研究结果显示,0.1、0.5、1、5、10 mg/mL的NMN均能促进线虫体长的增加,这与吴长泉[46]在NMN可促进小鼠毛发生长以及王柯诺[47]在NMN可促进酵母菌生长方面的研究结果相似,表明NMN处理可有效促进生长。此外,王丽鑫[48]研究发现采用人参挥发油喂食线虫8 d,线虫的体长相较对照组增加了13.25%。金司仪等[49]报道仅三七细粉高剂量组(40 mg/mL)能增加线虫不到15%的体长,而其他剂量组差异均无统计学意义。童杰文[50]指出红茶提取物和绿茶提取物可以分别增加线虫11.38%、7.09%的体长。Zhang等[51]研究表明紫甘蓝汁和绿甘蓝汁可以分别增加线虫12.52%、10.42%的体长。而本研究在最佳质量浓度1 mg/mL NMN喂食的情况下,线虫的体长在第4、7、10、13 d相较其对应的空白对照组分别提高可达62.82%、54.02%、25.46%、35.24%,体现出了NMN促生长应用的优势和潜在价值。
秀丽隐杆线虫的生长发育由很多基因决定,其中胶原蛋白作为一类关键的结构蛋白在线虫的生长发育过程中起着重要作用[52]。秀丽隐杆线虫有175个胶原蛋白,主要分为外皮胶原蛋白和基底膜胶原蛋白两大类,与哺乳动物具有较高的同源性[53]。研究发现一些胶原蛋白基因的突变或沉默可以显著影响线虫的生长发育,如col-121、dpy-5基因表达沉默的线虫身体更为短粗或细长,dpy-2、dpy-7、dpy-10、dpy-13、rol-6、sqt-1和sqt-3等基因的突变会引起线虫变短或表皮起泡等[54-55]。本研究采用转录组测序技术解析NMN对线虫促生长的作用机制,检测到喂食NMN可以显著改变线虫基因的表达水平,而筛选出的差异表达基因主要富集在胶原蛋白参与的角质层发育和基于角质素的角质层蜕皮周期、基于胶原蛋白和角质素的角质层发育、胶原蛋白三聚体等GO条目,这些属于生物过程类别的GO条目均与生长发育有关,结果表明NMN可能通过改变这些GO条目中基因的表达促进线虫的生长发育。进一步基于GO富集分析和差异表达基因蛋白互作网络分析筛选出可能参与NMN促线虫生长过程的关键核心基因,包括col-107、dpy-13、col-125、dpy-4、col-180、col-168、col-167、sqt-1、col-94、col-144、等。值得一提的是,线虫在自然生长发育的过程中,一些与角质层发育和蜕皮相关的基因家族,如dpy、col、sqt、lon、rol等会出现表达差异[56]。此外,dpy-13、dpy-7、dpy-4、sqt-1、rol-6已被证实可以影响线虫的生长发育[44-45,52,57],这些研究提示NMN可能通过调控这些差异表达基因从而促进线虫的生长。
综上所述,NMN具有较强的促生长功效,能显著增加线虫的体长,其原因可能是喂食NMN后,线虫生长过程中一些与胶原蛋白和角质层发育有关的生物学功能及相关基因受到调控,其中col-107、dpy-13、col-125、dpy-4、col-180、col-168、col-167、sqt-1、col-94、col-144等可能是关键的差异表达基因。本研究拓宽了NMN的应用范围,为开发促生长功能性食品、保健品、药品或饲料等提供了参考。未来仍需深入验证转录组学的推测结果,进一步开展相关差异表达基因的功能研究。
-
![]()
图 1 NMN喂食线虫第13 d的照片
注:A、B、C、D、E、F分别为空白组、0.1、0.5、1、5、10 mg/mL NMN处理组。
Figure 1. Photograph of C. elegans on day 13 of NMN treatment
![]()
图 2 差异表达基因分析与筛选
注:A、B、C、D分别为A07vsAN7、A013vsAN13、A00vsA07、A07vsA013差异表达基因火山图;E为A07vsAN7和A013vsAN13的共同上调差异基因韦恩图;F为A00vsA07和A07vsA013的共同上调差异基因韦恩图。
Figure 2. Analysis and screening of differentially expressed genes
![]()
图 3 差异表达基因GO富集分析
注:图中圆点的颜色对应不同的P值范围;圆点的大小代表在该GO条目中的基因数量;富集因子表示该GO条目中富集到的基因数量占该GO条目中总基因数量的比值,富集因子越大,表示GO的富集程度越大。
Figure 3. GO enrichment analysis of differentially expressed genes
![]()
图 4 差异表达基因GO有向无环图
Figure 4. GO directed acyclic graph of differentially expressed genes
![]()
图 5 差异表达基因蛋白互作网络图
注:图中Degree值代表节点的连通性,节点的颜色对应不同的Degree值范围,Degree值越大,说明与该节点相互作用的基因越多;节点大小代表基因在该网络中的重要程度,节点越大,说明基因在该网络中越重要。
Figure 5. Protein-protein interaction network of differentially expressed genes
![]()
图 6 A07vsAN7(A)和A013vsAN13(B)关键核心基因差异表达验证
Figure 6. Validation of differential expression of key genes in A07vsAN7 (A) and A013vsAN13 (B)
表 1 关键核心基因引物序列
Table 1 Primer sequences of key genes
基因 上游引物(5’-3’) 下游引物(5’-3’) col-107 AGCCATCGTTATTGGAGCGTCTTTC AGGCTTCATTGGCGGTGTTTCTG dpy-13 CACAGAAGCCTTGCGAGGAGATC TCCTGGCTGTCCTGGTTGTCC col-125 GAAACCGTACCGCCCGTCAAG TGGAAGGCAGCAAGCATCACATC dpy-4 AACCGTACCGCCCGTTCTACC GCTCCTGGCTTTCCTGGCTTTC col-180 CCGCACTCAAGTTCTCGCTAAGG CAGCAGCAGAGACACCGCATC col-168 TCTCCTTCACTGCCACTTTG TCCAGCAGCAGATACTCCAC col-167 CGTCTCCTTCACTGCCACA ACTCCACATCCGCAGCAT sqt-1 AGCGTGTCCGTCGTCAATATGAAG CTGGTGGAACTGCTGGTGGTTG col-94 CTTCCCGTTTCGCCCGTCAAG GCATCCAGAGCATGATCCTCCTTG col-144 TGCCAATGCGAGCCAACTAAGC AGGTGAGTGGAGCGAAGGTAGC act-1 TCGGTATGGGACAGAAGGAC CATCCCAGTTGGTGACGATA  下载: 导出CSV
下载: 导出CSV
表 2 NMN对线虫体长的影响
Table 2 Effect of NMN on the body length of C. elegans
组别 平均体长(μm) 第4 d 第7 d 第10 d 第13 d 空白组 1097.21±58.30 1358.12±226.87 1744.96±179.89 1724.14±197.68 0.1 mg/mL NMN 1690.08±127.68**** 1874.49±175.45*** 2010.70±115.03* 1920.16±130.65* 0.5 mg/mL NMN 1682.49±73.88**** 1845.79±246.27*** 2121.82±138.26*** 2124.91±149.15**** 1 mg/mL NMN 1786.50±74.23**** 2091.75±62.33**** 2189.21±165.56**** 2331.77±94.70**** 5 mg/mL NMN 1779.99±140.23**** 1994.70±221.00**** 2133.55±252.82**** 2127.99±113.93**** 10 mg/mL NMN 1580.63±118.64**** 2048.12±206.78**** 2176.72±148.28**** 2112.67±165.67**** 注:*与空白组相比,P<0.05;***与空白组相比,P<0.001;****与空白组相比,P<0.0001。
下载: 导出CSV
表 3 线虫样本转录组测序统计
Table 3 Transcriptomic sequencing statistics of C. elegans
样本编号 Raw reads Clean reads Q20(%) Q30(%) 比对率(%) A001 59049504 57669184 97.31 92.69 95.90 A002 44952038 44653322 97.68 93.36 95.90 A003 53516110 52311272 97.21 92.39 95.97 A071 55185606 53795278 97.47 93.01 96.48 A072 43503612 43215388 97.76 93.57 95.90 A073 56342184 55299230 97.77 93.68 96.49 AN71 54934544 54178138 97.77 93.72 96.46 AN72 57107824 56171120 97.29 92.52 95.98 AN73 61027166 59699564 97.61 93.35 96.20 A0131 44940238 44590662 97.40 92.71 95.56 A0132 55203858 54770834 97.51 92.96 95.78 A0133 64016058 63209158 97.54 93.07 96.08 AN131 44595066 44277946 97.69 93.41 95.61 AN132 59915312 58374528 97.37 92.74 96.30 AN133 56551810 56066104 97.66 93.33 95.85 注:样本编号的最后1位数字1、2、3分别代表该组的3个平行样本。
下载: 导出CSV
-
[1] COVARRUBIAS A J, PERRONE R, GROZIO A, et al. NAD+ metabolism and its roles in cellular processes during ageing[J]. Nature Reviews Molecular Cell Biology,2021,22(2):119−141. doi: 10.1038/s41580-020-00313-x
[2] BERTOLDO M J, LISTIJONO D R, HO W J, et al. NAD+ repletion rescues female fertility during reproductive aging[J]. Cell Reports,2020,30(6):1670−1681. doi: 10.1016/j.celrep.2020.01.058
[3] VERDIN E. NAD+ in aging, metabolism, and neurodegeneration[J]. Science,2015,350(6265):1208−1213. doi: 10.1126/science.aac4854
[4] KINCAID J W, BERGER N A. NAD+ metabolism in aging and cancer[J]. Experimental Biology and Medicine,2020,245(17):1594−1614. doi: 10.1177/1535370220929287
[5] BRAIDY N, LIU Y. NAD+ therapy in age-related degenerative disorders: A benefit/risk analysis[J]. Experimental Gerontology,2020,132:110831. doi: 10.1016/j.exger.2020.110831
[6] HONG W, MO F, ZHANG Z, et al. Nicotinamide mononucleotide: A promising molecule for therapy of diverse diseases by targeting NAD+ metabolism[J]. Frontiers in Cell and Developmental Biology,2020,8:246. doi: 10.3389/fcell.2020.00246
[7] CROTEAU D L, FANG E F, NILSEN H, et al. NAD+ in DNA repair and mitochondrial maintenance[J]. Cell Cycle,2017,16(6):491−492. doi: 10.1080/15384101.2017.1285631
[8] RU M, WANG W, ZHAI Z, et al. Nicotinamide mononucleotide supplementation protects the intestinal function in aging mice and D-galactose induced senescent cells[J]. Food Function,2022,13(14):7507−7519. doi: 10.1039/D2FO00525E
[9] KIM M, SEOL J, SATO T, et al. Effect of 12-week intake of nicotinamide mononucleotide on sleep quality, fatigue, and physical performance in older Japanese adults: A randomized, double-blind placebo-controlled study[J]. Nutrients,2022,14(4):755. doi: 10.3390/nu14040755
[10] BRITO S, BAEK J M, CHA B, et al. Nicotinamide mononucleotide reduces melanin production in aged melanocytes by inhibiting cAMP/Wnt signaling[J]. Journal of Dermatological Science,2022,106(3):159−169. doi: 10.1016/j.jdermsci.2022.05.002
[11] JIANG Y, DENG Y, PANG H, et al. Treatment of SARS-CoV-2-induced pneumonia with NAD+ and NMN in two mouse models[J]. Cell Discovery,2022,8(1):38. doi: 10.1038/s41421-022-00409-y
[12] HONG S M, LEE A Y, HWANG S M, et al. NAMPT mitigates colitis severity by supporting redox-sensitive activation of phagocytosis in inflammatory macrophages[J]. Redox Biology,2022,50:102237. doi: 10.1016/j.redox.2022.102237
[13] KIM H W, RYOO G H, JANG H Y, et al. NAD+-boosting molecules suppress mast cell degranulation and anaphylactic responses in mice[J]. Theranostics,2022,12(7):3316−3328. doi: 10.7150/thno.69684
[14] NAGAHISA T, YAMAGUCHI S, KOSUGI S, et al. Intestinal epithelial NAD+ biosynthesis regulates GLP-1 production and postprandial glucose metabolism in mice[J]. Endocrinology,2022,163(4):bqac023. doi: 10.1210/endocr/bqac023
[15] BAKER B L, NEECE C L, FENNING R M, et al. Mental disorders in five-year-old children with or without developmental delay: Focus on ADHD[J]. Journal of Clinical Child and Adolescent Psychology,2010,39(4):492−505. doi: 10.1080/15374416.2010.486321
[16] MARQUIS W A, BAKER B L. An examination of Anglo and Latino parenting practices: Relation to behavior problems in children with or without developmental delay[J]. Research in Developmental Disabilities,2014,35(2):383−392. doi: 10.1016/j.ridd.2013.11.010
[17] BRIAUX J, MARTIN-PREVEL Y, CARLES S, et al. Evaluation of an unconditional cash transfer program targeting children's first-1, 000-days linear growth in rural Togo: A cluster-randomized controlled trial[J]. PLoS Medicine,2020,17(11):e1003388. doi: 10.1371/journal.pmed.1003388
[18] 王凤, 肖楚翔, 刘淑珍, 等. 榴莲核黄酮的提取及其对秀丽隐杆线虫氧化和衰老的影响[J]. 食品科学,2021,42(9):123−129. [WANG F, XIAO C X, LIU S Z, et al. Extraction of flavonoids from durian seeds and its antioxidant and anti-aging effects in Caenorhabditis elegans[J]. Food Science,2021,42(9):123−129. WANG F, XIAO C X, LIU S Z, et al. Extraction of flavonoids from durian seeds and its antioxidant and anti-aging effects in Caenorhabditis elegans[J]. Food Science, 2021, 42(9): 123-129.
[19] 沈静德. 秀丽隐杆线虫是研究发育生物学的好材料[J]. 生物学通报,1991(9):16−18. [SHEN J D. Caenorhabditis elegans is a good material for studying developmental biology[J]. Bulletin of Biology,1991(9):16−18. SHEN J D. Caenorhabditis elegans is a good material for studying developmental biology[J]. Bulletin of biology, 1991(9): 16-18.
[20] 夏程程, 杨番, 钟晓凌, 等. 秀丽隐杆线虫模型在食品安全研究中的应用[J]. 中国食品学报,2020,20(6):312−323. [XIA C C, YANG F, ZHONG X L, et al. Application of Caenorhabditis elegans model in food safety research[J]. Journal of Chinese Institute of Food Science and Technology,2020,20(6):312−323. XIA C C, YANG F, ZHONG X L, et al. Application of Caenorhabditis elegans model in food safety research[J]. Journal of Chinese Institute of Food Science and Technology, 2020, 20(6): 312-323.
[21] 程雷. 稳态磁场对秀丽隐杆线虫神经行为和早期胚胎发育的影响[D]. 合肥: 中国科学技术大学, 2022: 81−82 CHEN L. Effect of static magnetic fields on neurobehavior and early embryonic development in Caenorhabditis elegans[D]. Hefei: University of Science and Technology of China, 2022: 81−82.
[22] PRADHAN A, OLSSON P E, JASS J. Di(2-ethylhexyl) phthalate and diethyl phthalate disrupt lipid metabolism, reduce fecundity and shortens lifespan of Caenorhabditis elegans[J]. Chemosphere,2018,190(1):375−382.
[23] 石珏, 杨亚宁, 黎青青, 等. 双酚A不同暴露方式对线虫生长生殖发育及凋亡的影响[J]. 环境化学,2018,37(6):1203−1208. [SHI J, YANG Y N, LI Q Q, et al. Effects of bisphenol A on the growth, reproductive development and apoptosis in Caenorhabditis elegans under different exposure routes[J]. Environmental Chemistry,2018,37(6):1203−1208. SHI J, YANG Y N, LI Q Q, et al. Effects of bisphenol A on the growth, reproductive development and apoptosis in Caenorhabditis elegans under different exposure routes[J]. Environmental Chemistry, 2018, 37(6): 1203-1208.
[24] 叶丽云, 程冰, 马水丽, 等. 赤芝多糖对小鼠急性酒精性肝损伤的保护效果和作用机制[J]. 食品科学,2022,43(5):103−110. [TE L Y, CHENG B, MA S L, et al. Preventive effect and mechanism of Ganoderma lingzhi polysaccharides on acute alcoholic liver injury in mice[J]. Food Science,2022,43(5):103−110. TE L Y, CHENG B, MA S L, et al. Preventive effect and mechanism of Ganoderma lingzhi polysaccharides on acute alcoholic liver injury in mice[J]. Food Science, 2022, 43(5): 103-110.
[25] 何秋玲, 张彩平, 桂静, 等. 异麦芽酮糖减少小鼠肝脏脂肪堆积的关键基因筛选与验证[J]. 食品工业科技,2022,43(16):1−8. [HE Q L, ZHANG C P, GUI J, et al. Screening and verification of key genes for isomaltulose reducing liver fat accumulation in mice[J]. Science and Technology of Food Industry,2022,43(16):1−8. HE Q L, ZHANG C P, GUI J, et al. Screening and verification of key genes for isomaltulose reducing liver fat accumulation in mice[J]. Science and Technology of Food Industry, 2022, 43(16): 1-8.
[26] 张婷婷, 苗玥, 吴恩凯, 等. 普洱茶茶褐素对代谢综合征大鼠生长过程中肝脏差异表达基因的影响[J]. 食品工业科技,2022,43(7):1−9. [ZHANG T T, MIAO Y, WU E K, et al. Effects of theabrownin from puerh tea on differentially expressed genes in the liver of MS rats during growth[J]. Science and Technology of Food Industry,2022,43(7):1−9. ZHANG T T, MIAO Y, WU E K, et al. Effects of theabrownin from puerh tea on differentially expressed genes in the liver of MS rats during growth[J]. Science and Technology of Food Industry, 2022, 43(7): 1-9.
[27] 丁明孝, 苏都莫日根, 王喜忠, 等. 细胞生物学实验指南, 第二版[M]. 北京: 高等教育出版社, 2013: 173−175 DING M X, SDMRG, WANG X Z, et al. Experiments in cell biology, Second edition[M]. Beijing: Higher education press, 2013: 173−175.
[28] DEHGHAN E, ZHANG Y, SAREMI B, et al. Hydralazine induces stress resistance and extends C. elegans lifespan by activating the NRF2/SKN-1 signalling pathway[J]. Nature Communications,2017,8(1):2223. doi: 10.1038/s41467-017-02394-3
[29] LI P, WANG Z, LAM S M, et al. Rebaudioside a enhances resistance to oxidative stress and extends lifespan and healthspan in Caenorhabditis elegans[J]. Antioxidants,2021,10(2):262. doi: 10.3390/antiox10020262
[30] 檀克勤. 基于秀丽隐杆线虫筛选抗大肠杆菌K88感染的乳酸菌及其作用机制[D]. 广州: 暨南大学, 2019: 60−61 TAN K Q. Screening of lactic acid bacteria against Escherichia coli K88 infection based on Caenorhabditis elegans and its mechanism of action[D]. Guangzhou: Jinan University, 2019: 60−61.
[31] 许吉安. 铁皮石斛中性多糖对秀丽隐杆线虫寿命的影响[D]. 广州: 华南农业大学, 2018: 23−24 XU J A. Effect of Dendrobium officinale polysaccharides on the lifespan of Caenorhabditis elegans and its mechanism[D]. Guangzhou: South China Agricultural University, 2018: 23−24.
[32] KIM D, LANGMEAD B, SALZBERG S L. HISAT: A fast spliced aligner with low memory requirements[J]. Nature Methods,2015,12(4):357−360. doi: 10.1038/nmeth.3317
[33] LOVE M, HUBER W, ANDERS S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2[J]. Genome Biology,2014,15(12):550. doi: 10.1186/s13059-014-0550-8
[34] ZHENG Q, WANG X J. GOEAST: A web-based software toolkit for gene ontology enrichment analysis[J]. Nucleic Acids Research,2008,36(Suppl2):W358−W363.
[35] 白娟, 张金富, 张佩熙, 等. 苦瓜皂苷对秀丽隐杆线虫寿命的影响及其机制研究[J]. 食品科学,2022,43(7):165−173. [BAI J, ZHANG J F, ZHANG P X, et al. Effect of saponin from bitter melon (Momordica charantia) on the lifespan of Caenorhabditis elegans and its mechanism[J]. Food Science,2022,43(7):165−173. BAI J, ZHANG J F, ZHANG P X, et al. Effect of saponin from bitter melon (Momordica charantia) on the lifespan of Caenorhabditis elegans and its mechanism[J]. Food Science, 2022, 43(7): 165-173.
[36] 王晨. 磷酸三(1, 3-二氯-2-丙基)酯诱导秀丽隐杆线虫衰老效应及健康风险分子机制[D]. 上海: 华东理工大学, 2020: 58 WANG C. Molecular mechanisms of aging effects of C. elegans and health risks induced by Tris (1, 3-dichloro-2-propyl) phosphate[D]. Shanghai: East China University of Science and Technology, 2020: 58.
[37] 李博. 藻毒素BMAA在土壤与作物间的迁移累积及其对秀丽隐杆线虫的影响[D]. 南京: 南京农业大学, 2019: 40 LI B. Transfer and bioaccumulation of a cyanobacterial neurotoxin BMAA between soil and crop and its effects on Caenorhabditis elegans[D]. Nanjing: Nanjing Agricultural University, 2019: 40.
[38] 侯殿志, 唐健, 郑博妍, 等. 基于转录组学的绿豆改善肥胖小鼠肝脂肪变性的潜在机制[J]. 食品科学技术学报,2021,39(6):45−52. [HOU D Z, TANG J, ZHENG B Y, et al. Potential mechanisms of mung bean in improvement of hepatic steatosis in obese mice based on transcriptomics[J]. Journal of Food Science and Technology,2021,39(6):45−52. HOU D Z, TANG J, ZHENG B Y, et al. Potential mechanisms of mung bean in improvement of hepatic steatosis in obese mice based on transcriptomics[J]. Journal of Food Science and Technology, 2021, 39(6): 45-52.
[39] 丛岩懿. 线虫酸性pH胁迫响应机制的初步研究[D]. 北京: 中国科学院大学, 2020: 48 CONG Y Y. A preliminary study on acidic pH stress responses mechanisms of nematodes[D]. Beijing: University of Chinese Academy of Sciences, 2020: 48.
[40] 杨文清. 吩嗪通过组蛋白去乙酰化酶HAD-1抑制线虫天然免疫分子机制的研究[D]. 昆明: 云南大学, 2020: 44 YANG W Q. Molecular mechanism of phenazine inhibits innate immunity of C. elegans by histone deacetylase HAD-1[D]. Kunming: Yunnan University, 2020: 44.
[41] 陈媛丽. 运动延长秀丽隐杆线虫寿命的分子机制研究[D]. 昆明: 云南大学, 2019: 61 CHEN Y L. The mechanism of lifespan extension induced by exercise in C. elegans[D]. Kunming: Yunnan University, 2019: 61.
[42] 杨佳彤. 北细辛对秀丽隐杆线虫生物毒性作用研究[D]. 长春: 吉林农业大学, 2022: 54 YANG J T. Study on the biological toxicity of Asarum heterotropoides to Caenorhabditis elegans[D]. Changchun: Jilin Agricultural University, 2022: 54.
[43] 史强. 基于秀丽隐杆线虫模型研究卵转铁蛋白对机体的影响及作用机制初探[D]. 南昌: 南昌大学, 2022: 51 SHI Q. Effects of egg ovotransferrin on Caenorhabditis elegans and its action mechanism[D]. Nanchang: Nanchang University, 2022: 51.
[44] MENDE N V, BIRD D M, ALBERT P S, et al. dpy-13: a nematode collagen gene that affects body shape[J]. Cell,1988,55(4):567−576. doi: 10.1016/0092-8674(88)90215-2
[45] KRAMER J M, JOHNSON J J, EDGAR R S, et al. The sqt-1 gene of C. elegans encodes a collagen critical for organismal morphogenesis[J]. Cell,1988,55(4):555−565. doi: 10.1016/0092-8674(88)90214-0
[46] 吴长泉. β-烟酰胺单核苷酸在预防脱发或促进毛发生长产品中的应用: CN 202010549459.2[P] 2020-06-16]. [WU C Q. β-Application of nicotinamide mononucleotide in hair loss prevention or hair growth promotion products: CN 202010549459.2[P]. [2020-06-16
[47] 王柯诺. 三类功效成分对真核生物生长和衰老的影响[D]. 杭州: 浙江工商大学, 2021: 18 WANG K N. Effects of three functional components on growth and senescence of eukaryotes[D]. Hangzhou: Zhejiang Gongshang University, 2021: 18.
[48] 王丽鑫. 人参挥发油延长秀丽隐杆线虫寿命及健康寿命的研究[D]. 长春: 吉林大学, 2022: 32 WANG L X. The study on ginseng volatile oil prolongs the lifespan and healthspan of Caenorhabditis elegans[D]. Changchun: Jilin University, 2022: 32.
[49] 金司仪, 洪怡, 卢山, 等. 不同粒度三七醇提物对秀丽隐杆线虫毒性作用的研究[J]. 毒理学杂志,2019,33(3):208−212. [JIN S Y, HONG Y, LU S, et al. Study on toxicity of alcohol extracts of different sizes of notoginseng to C. elegans[J]. Journal of Toxicology,2019,33(3):208−212. doi: 10.16421/j.cnki.1002-3127.2019.03.018 JIN S Y, HONG Y, LU S, et al. Study on toxicity of alcohol extracts of different sizes of Notoginseng to C. elegans[J]. Journal of Toxicology, 2019, 33(3): 208-212. doi: 10.16421/j.cnki.1002-3127.2019.03.018
[50] 童杰文. 红茶改善秀丽隐杆线虫紫外辐射应激作用与机制研究[D]. 长沙: 湖南农业大学, 2017: 17 TONG W J. Black tea improves resistance of Caenorhabditis elegans under radiation stress of ultraviolet[D]. Changsha: Hunan Agricultural University, 2017: 17.
[51] ZHANG N, JIAO S, JING P. Red cabbage rather than green cabbage increases stress resistance and extends the lifespan of Caenorhabditis elegans[J]. Antioxidants,2021,10(6):930. doi: 10.3390/antiox10060930
[52] 黄贝贝. 秀丽隐杆线虫(Caenorhabditis elegans)col-119基因的表达及功能分析[D]. 武汉: 华中师范大学, 2015: 5−6 HUANG B B. Expression and functional analysis to the gene col-119 of Caenorhabditis elegans[D]. Wuhan: Central China Normal University, 2015: I, 5−6.
[53] 李文娜. 胶原蛋白网络对秀丽隐杆线虫表皮固有免疫的调节的研究[D]. 苏州: 苏州大学, 2016 LI W N. A collagen network regulates innate immunity in C. elegans epidermis[D]. Suzhou: Soochow University, 2016.
[54] 逯召莲. 胶原蛋白基因col-131、col-150、col-121和dpy-5对秀丽隐杆线虫生殖与发育的影响[D]. 兰州: 兰州大学, 2015: 4 LU Z L. The effects of collagen genes col-131, col-150, col-121 and dpy-5 on fertility and development of Caenorhabditis elegans[D]. Lanzhou: Lanzhou University, 2015: 4.
[55] JOHNSTONE I L. Cuticle collagen genes[J]. Trends in Genetics,2000,16(1):21−27. doi: 10.1016/S0168-9525(99)01857-0
[56] GAO Y, LU Y, YI J, et al. A genome-wide mRNA expression profile in Caenorhabditis elegans under prolonged exposure to 1750 MHz radiofrequency fields[J]. Plos One,2016,11(1):e0147273. doi: 10.1371/journal.pone.0147273
[57] PARK Y S, KRAMER J M. The C. elegans sqt-1 and rol-6 collagen genes are coordinately expressed during development, but not at all stages that display mutant phenotypes[J]. Developmental Biology,1994,163(1):112−124. doi: 10.1006/dbio.1994.1127
 下载:
下载:










计量
- 文章访问数: 163
- HTML全文浏览量: 89
- PDF下载量: 21
