


Analysis of Codon Usage Bias in the Genome of Trichoderma reesei
-
摘要: 在不同物种中,密码子偏好性存在一定程度的差异。为了研究纤维素酶主要工业生产菌株——里氏木霉(Trichoderma reesei)基因组密码子的偏好性,对里氏木霉9352个基因的编码区进行密码子分析。结果显示,里氏木霉97%的基因GC含量为50%~68%,GC3的平均含量为70.4%。中性分析与ENC-plot分析表明,里氏木霉密码子的使用主要受选择压力的影响。相关性分析结果显示,基因组GC含量与GC1、GC2和GC3显著相关(P<0.05),有效密码子数与GC3显著相关(P<0.05)。此外,在里氏木霉使用频率较高的24个密码子中,有22个均是以GC结尾的。进一步确定了21个高表达优越密码子和4个高表达最优密码子(CUC、GCC、CGC和GGC)。里氏木霉与长梗木霉、粗糙脉孢霉在密码子使用频率上差异较小,与酿酒酵母的差异相对较大。本研究为里氏木霉中的密码子优化提供了理论依据,对开发高效的里氏木霉基因表达宿主以及开发里氏木霉作为合成生物学基盘细胞具有重要的意义。
-
关键词:
- 里氏木霉 /
- 密码子偏好性 /
- 有效密码子数 /
- 相对同义密码子使用度 /
- 异源表达
Abstract: Codon usage bias is quite different among different organisms. To elucidate the genetic codon preference in the most commonly used industrial strain Trichoderma reesei, codon usage analysis was performed with the opening reading frames of 9352 genes in T. reesei. The results showed that the GC content of 97% of T. reesei genes was 50%~68%, and the average content of GC3 was 70.4%. Neutral analysis and ENC-plot analysis demonstrated that the usage bias of T. reesei codons was mainly affected by selection pressure. Correlation analysis results revealed that the GC content of the genome was significantly correlated with GC1, GC2 and GC3 (P<0.05), and the number of effective codons was significantly correlated with GC3 (P<0.05). In addition, among the 24 codons frequently used by T. reesei, 22 of them contained G or C in their wobble position. Moreover, 21 codons associated with relatively effective gene expression and 4 codons relevant to optimal gene expression were further identified. The analysis also showed that the codon usage bias in T. reesei was similar to that in Trichoderma longibrachiatum and Neurospora crassa, while it was relatively different from that in Saccharomyces cerevisiae. The study herein would provide a feasible and advantageous method for genetic codon optimization in T. reesei, which would contribute to developing T. reesei as an excellent host for recombinant gene expression, as well as chassis for synthetic biology. -
遗传信息是由三联体密码子记载的。由于密码子的简并性,大多数氨基酸是由2~6种同义密码子编码。不同的物种编码同种氨基酸所利用的密码子种类与使用频率存在差别,这种现象称为密码子偏好性(Codon Usage Bias)[1]。许多因素影响各种生物体中密码子的使用,如自然选择(基因表达水平[2]、RNA丰度[3]、基因长度[4-5]、基因翻译起始信号和蛋白质结构[6])和突变压力(GC含量、突变频率和模式),以及随机遗传漂变等[7-9]。密码子使用模式的全基因组研究对理解基因组中分子组织的基本特征具有重要意义。迄今为止,对于遗传密码子偏好性的研究主要集中在一些模式物种,包括模式真菌酿酒酵母(Saccharomyces cerevisiae)、模式细菌大肠杆菌(Escherichia coli)、模式植物拟南芥(Arabidopsis thaliana)等方面[10-11],相比之下,对丝状真菌的研究相对较少。
丝状真菌里氏木霉作为生产纤维素酶和半纤维素酶的工业微生物,具有生长环境粗放、稳定性好、安全无毒、产酶效率高等优点。在食品加工工业中,用纤维素酶对农产品进行预处理,可以使植物组织膨化松软,减少农产品营养物质的损失。里氏木霉除了在食品工业中的作用外,还用于生物乙醇[12]和工业酶的生产,具有广泛的生物应用价值。迄今为止,大约有243种通过微生物发酵制造的市售酶产品,其中30种是使用里氏木霉作为宿主制成的,其中21种是重组产品,用于饲料和技术应用,包括纺织品、纸浆和纸张等[13-17],因此里氏木霉具有重要的研究价值。
目前,里氏木霉QM6a菌株的基因组已经完成测序[18],这为研究该丝状真菌的分子生物学提供了有利条件。本文以里氏木霉基因组为研究对象,通过对编码序列的核苷酸组成及密码子的偏好性进行分析,探究影响里氏木霉密码子使用偏差的因素。本研究结果有助于阐明该物种分子进化的机制,同时为通过密码子优化提高里氏木霉外源基因表达水平提供了理论依据。
1. 材料与方法
1.1 材料与仪器
里氏木霉QM6a基因组数据来自Joint Genome Institute(基因联合研究所,JGI http://genome.jgi.doe.gov/portal/)公共数据库,基因组项目编号为1184794;使用Galaxy生物信息学分析平台(https://usegalaxy.org/)中的Fasta Statistics对里氏木霉QM6a进行统计、Filter sequences by length对CDS进行过滤、cusp对密码子的GC含量进行分析;利用Python 3.9中的biopython-1.79模块对序列进行处理;利用CodonW 1.4.2软件对各个CDS密码子进行分析;利用Origin 9.0进行数据统计与作图。
1.2 实验方法
1.2.1 基因的选择
通过JGI数据库下载CDS序列,由于计算短序列的密码子数没有生物学意义[19],因此利用Galaxy的Filter sequences by length脚本过滤长度小于300 bp的CDS,收集最终序列(包含9352个CDS)用于进一步分析。
1.2.2 GC含量统计
利用Galaxy的cusp脚本统计分析各基因GC总含量以及密码子的第1、2和3位碱基为G或C的含量比例,分别记为GC、GC1、GC2和GC3。其中GC3对密码子使用偏好性具有重要影响。
1.2.3 中性绘图分析
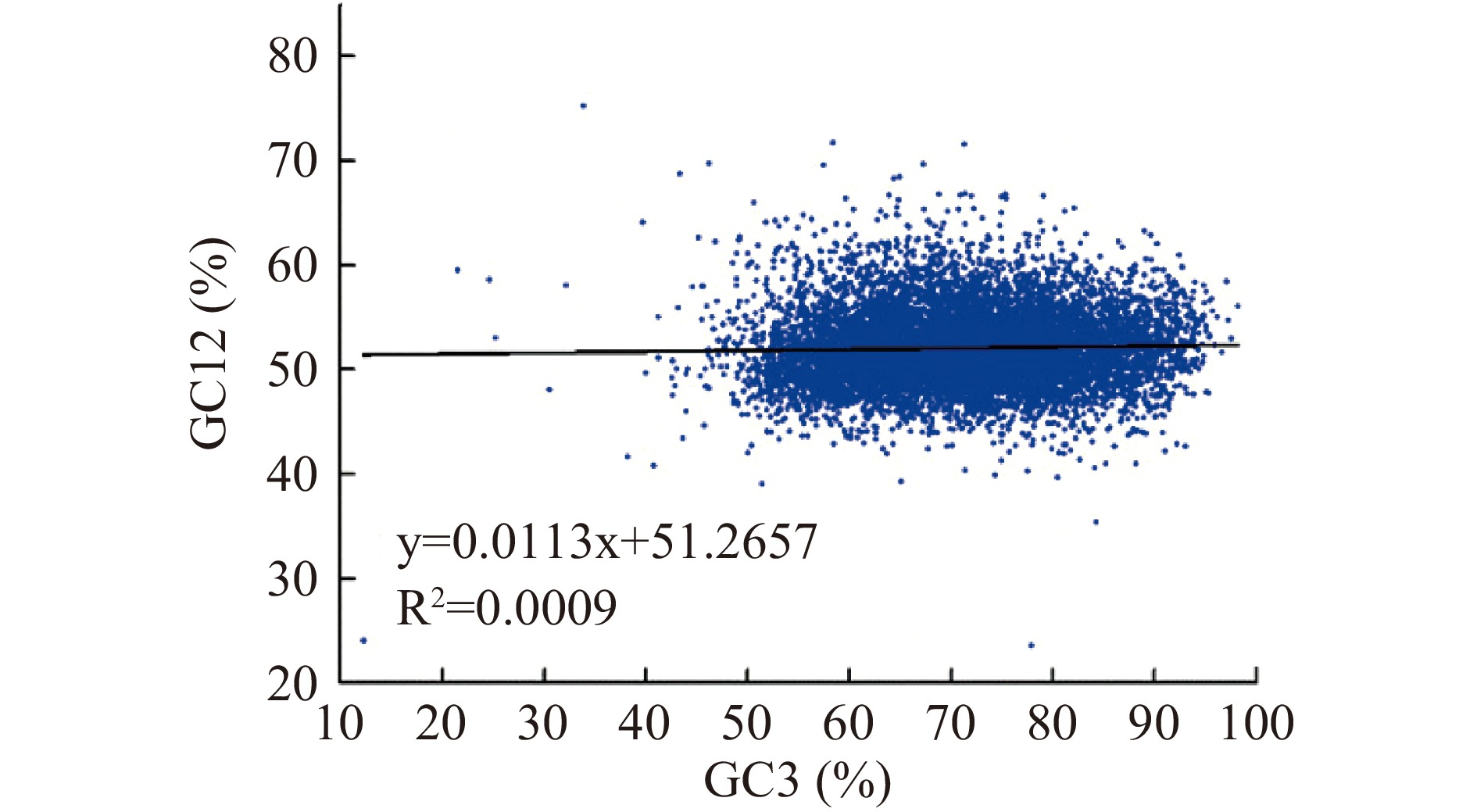
中性图是一种用于测量密码子使用模式的分析方法。本研究分析了第1、2和3位密码子位置(分别为GC1、GC2和GC3)的GC含量。GC12代表GC1和GC2的平均值;GC12和GC3用于中性绘图分析。在中性图中,如果GC12和GC3之间的相关性在统计学上显著,且回归线的斜率接近1,则假设突变偏差是影响密码子使用的主要因素。相反,针对突变偏倚的选择可能导致GC含量的窄分布以及GC12和GC3之间缺乏相关性[20]。
1.2.4 ENC-plot分析
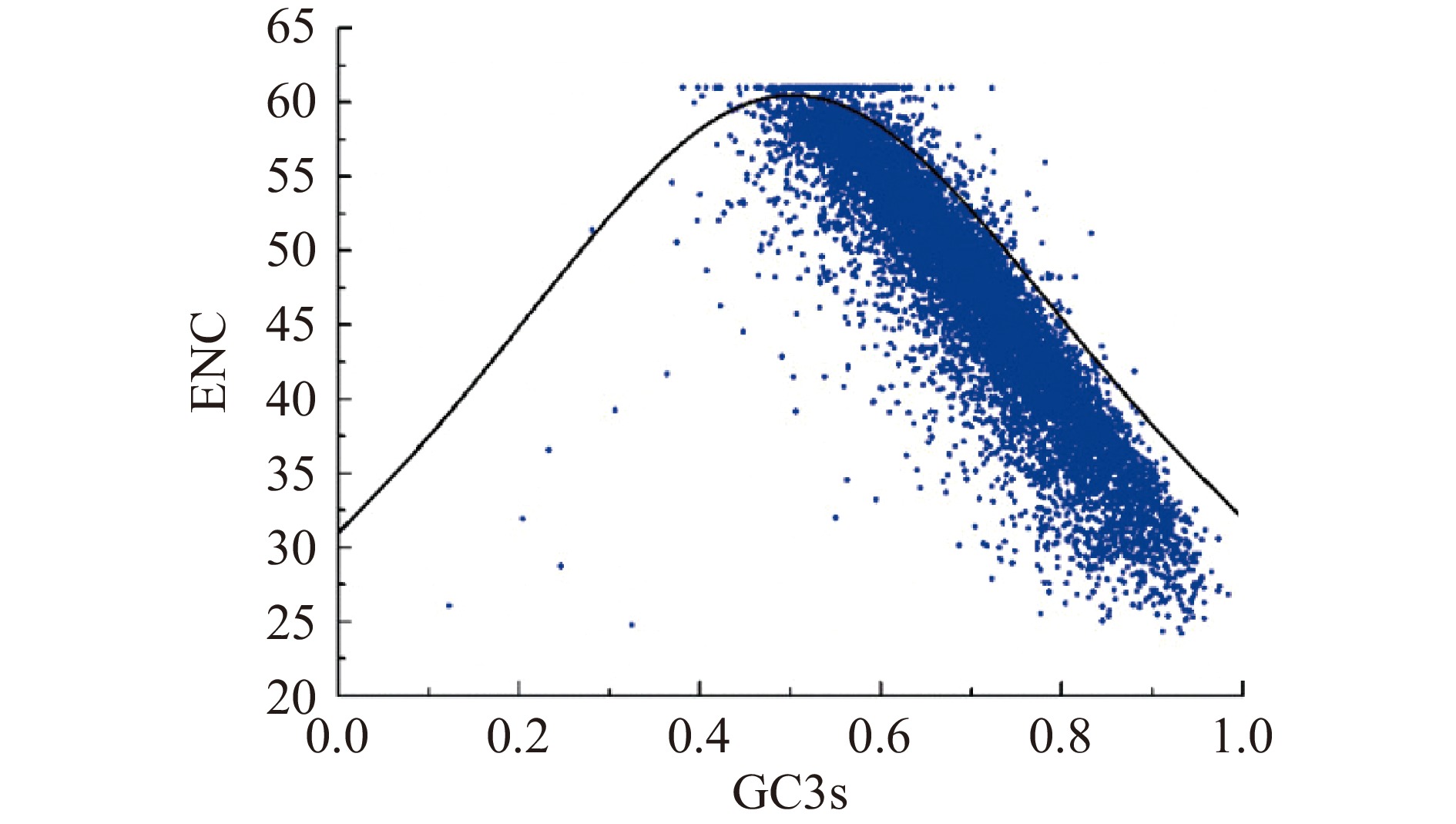
有效密码子数(Effective number of codon,ENC)提供了对绝对密码子偏差的有用估计,是确定某个基因的总体密码子使用偏差的一种度量。总GC含量,尤其是GC3(第三位的GC含量),经常反映定向突变的强度。以ENC为纵坐标,GC3为横坐标绘制的ENC-plot,广泛用于确定基因的密码子使用是否受到突变和选择的影响[21]。当对应点落在预期曲线附近时,突变是决定密码子使用的主要力量,当对应点大大低于预期曲线时,选择是决定密码子使用的主要力量。
1.2.5 相关性分析
变量和样本之间的关系可以通过多元统计分析来探索。使用皮尔森相关系数(Pearson correlation coefficient)进行相关性分析,使用双尾检测相关系数的显著性。相关性分析用来揭示密码子使用模式的主要因素并探究样本各变量之间的关联性[22]。
1.2.6 PR2-plot分析
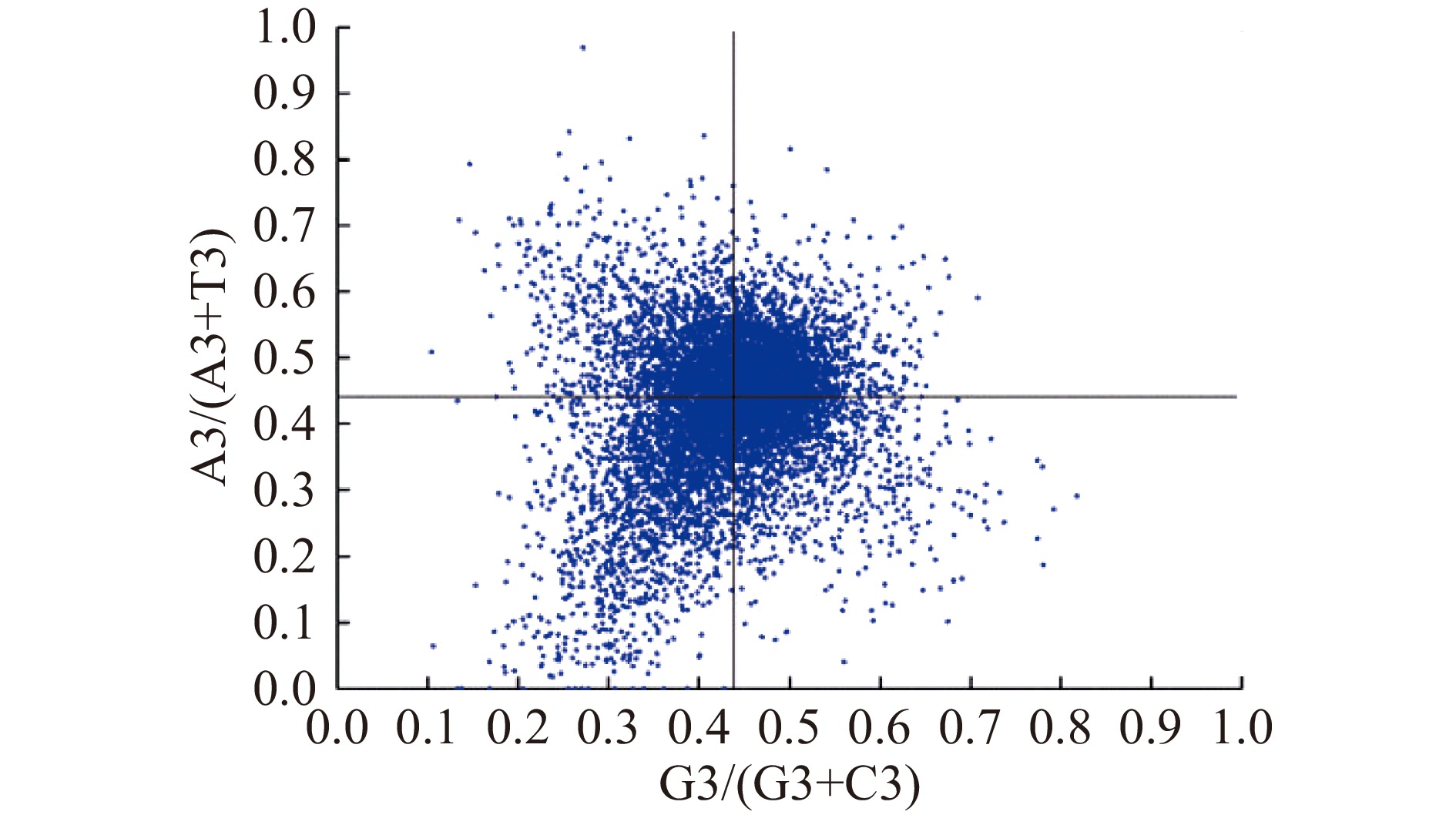
计算第3密码子位置(A3、U3、C3和G3)的核苷酸组成,并分析AT偏差(A3/(A3+U3))和GC偏差(G3/(G3+C3))。PR2-plot是以AT偏差(A3/(A3+T3))作为纵坐标和GC偏差(G3/(G3+C3))作为横坐标绘制的[23]。若核苷酸组成是影响同义密码子使用的唯一因素,那么A(T)和C(G)的使用频率应该相等。
1.2.7 相对同义密码子使用分析
运用CodonW软件分析相对同义密码子使用度(relative synonymous codon usage,RSCU)。RSCU是指对于某一特定的密码子在编码对应氨基酸的同义密码子的相对概率,它去除了氨基酸组成对密码子的影响。公式如下:
RSCUij=xij1nini∑j−1xij 式中:xij表示编码第i个氨基酸的第j个密码子的出现次数;ni总表示编码第i个氨基酸的同义密码子的数量(值为1~6)。如果密码子使用没有偏好,则该密码子的RSCU值等于1。当某一密码子的RSCU值大于1,则表明密码子的使用偏好性较强。由于它计算方便,而且很直观地反映出密码子使用的偏好性,因此在大多数的密码子相关分析中,都使用它作为衡量偏好性的标准。
1.2.8 高表达密码子的确定
ENC值的范围在20~61之间,单个基因的ENC值越低,该基因的整体密码子使用偏好就越强,基因的表达量相对越高[24]。故以ENC值为偏好性标准,两级各选10%的基因分别创建高低表达样本库,取两库△RSCU>0.08的密码子进行分析[25-26]。
1.2.9 与其他真菌密码子偏好性比较
运用Bioinformatics在线平台(http://www.bioinformatics.org)的Codon Usage计算里氏木霉各密码子的使用频率[27]。用CodonW分别计算出同属的长梗木霉(Trichoderma longibrachiatum)、丝状模式真菌粗糙脉孢霉(Neurospora crassa)、模式真菌酿酒酵母(Saccharomyces cerevisiae)的密码子使用频率,将里氏木霉密码子使用频率与它们进行比较分析。
2. 结果与分析
2.1 密码子偏性分析
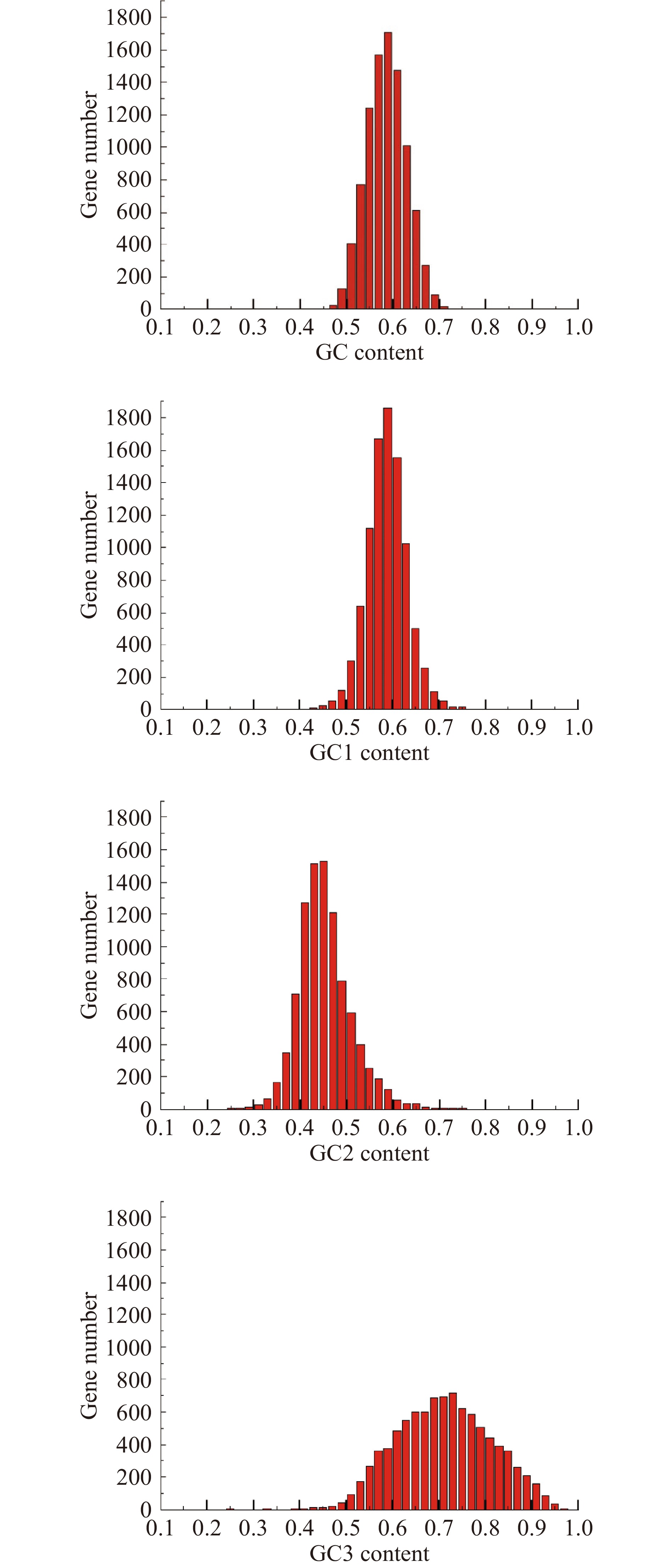
单一类型密码子的密码子使用偏差受基因组总核苷酸含量的影响很大[28],因此,首先利用Galaxy平台分析了里氏木霉基因组中编码序列(CDS)的GC核苷酸组成。在里氏木霉基因组中,97%的基因GC含量、96%的基因GC1含量、18%的基因GC2含量和37%的基因GC3含量分布在50%~68%(图1),GC核苷酸平均含量为58.1%,三个密码子位置(GC1、GC2和GC3)的GC平均含量分别为58.9%、45.0%和70.4%。单因素方差分析表明密码子三个位置的GC含量差异极显著(P<0.001),GC3>GC1>GC2表明第三位置的GC含量不同于第一和第二位置的GC含量,第三位密码子的GC含量在密码子位置中最高,说明GC3是导致密码子使用发生偏好性的重要原因,且里氏木霉的密码子第三位受到的选择压力较大。核苷酸组成分析结果表明,里氏木霉基因第三位偏好G/C末端密码子比A/U末端密码子更受欢迎。
2.2 中性绘图分析
中性分析是揭示GC12和GC3之间关系的一种有用方法。为了分析三个密码子位置之间的关系,本文构建了里氏木霉基因组编码序列的中性图(GC12与GC3)。结果显示(图2)GC12和GC3不相关(R2=0.0009),且斜率接近0,说明里氏木霉密码子没有受到定向突变压力的影响,导致密码子偏好性的原因主要是选择压力。
2.3 ENC-plot 分析
有效密码子数广泛用于测量单个基因的密码子偏好水平。为了阐明里氏木霉序列中核苷酸组成和密码子偏好之间的关系,绘制了ENC和GC3s图,从而探索了基因间密码子使用的主要特征。如图3所示,大部分基因的ENC观察值落在ENC期望值曲线之下,表明其里氏木霉密码子的使用主要受选择压力的影响,与中性绘图分析的结果一致。
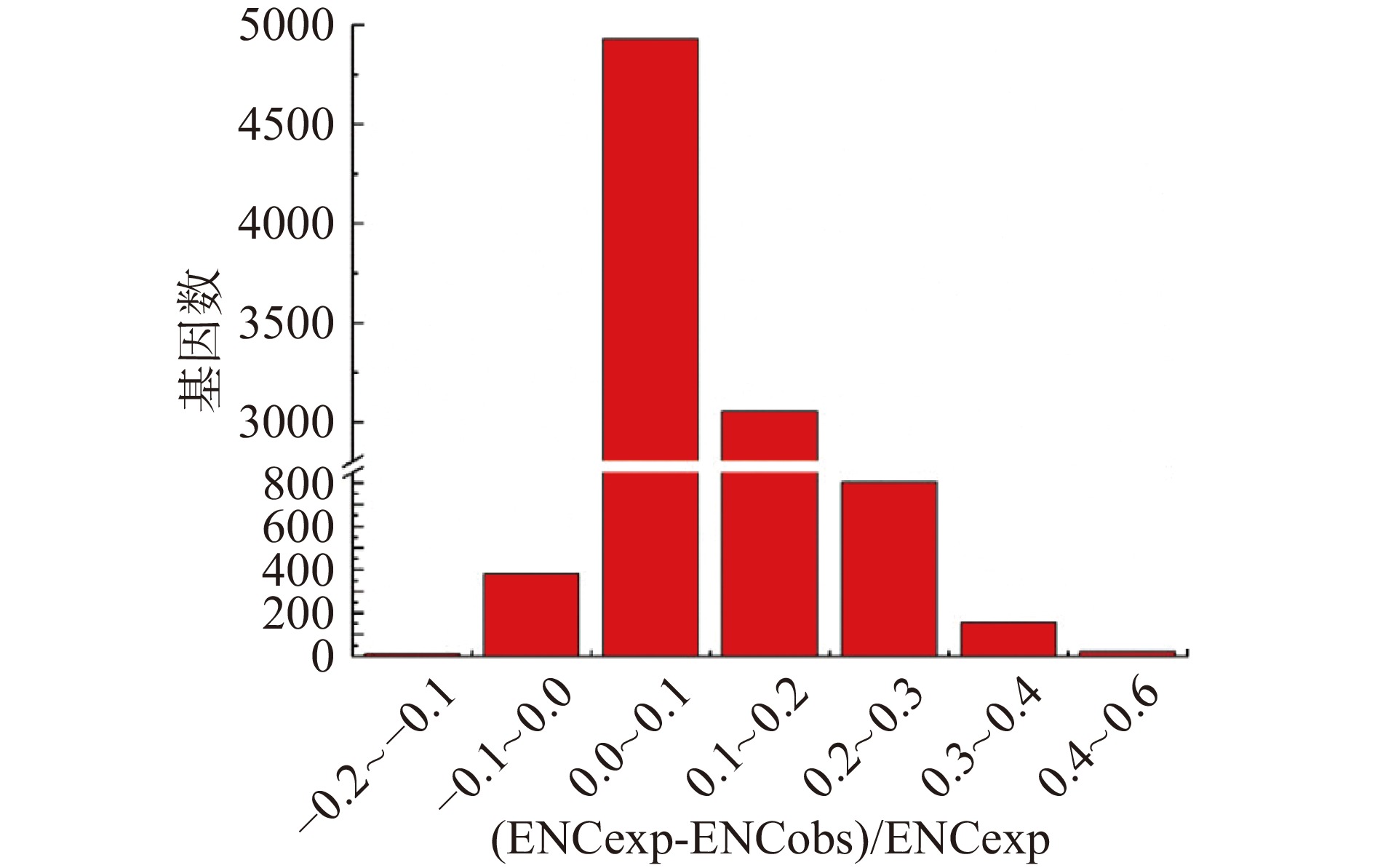
为了更准确地估计观测值和预期ENC值之间的差异,本文计算了(ENCexp-ENCobs)/ENCexp的值。如图4所示,(ENCexp-ENCobs)/ENCexp的峰值为0~0.1,表明大多数基因的ENC值与基于GC3的预期ENC值略有不同。因此,大多数基因观察到的ENC接近基于GC3的预期ENC,尽管有部分基因观察到的ENC要低得多。
![]() 图 4 有效密码子数(ENC)比率的频率分布图Figure 4. Frequency distribution of the effective number of codons (ENC) ratio
图 4 有效密码子数(ENC)比率的频率分布图Figure 4. Frequency distribution of the effective number of codons (ENC) ratio2.4 相关性分析
里氏木霉基因组编码区中GC含量、ENC和密码子出现频数(CN)间相关性分析结果显示(表1),GC Total与GC1、GC2、GC3呈极显著相关(P<0.001),GC3与GC1、GC2相关性水平不显著,说明GC3与GC1、GC2的密码子组成存在较大差异。进一步发现,ENC与GC1、GC2相关性较弱,与GC3以及GC TOTAL极显著相关(P<0.001),表明密码子不同位置的碱基组成会影响有效密码子数。CN值与GC1、GC2、GC3、GC Total相关性都不显著,说明CN对ENC的影响很小,排除了基因序列过短对后续分析的影响。
表 1 各基因相关参数的相关性分析Table 1. Correlation analysis of each gene-related parameters指标 GC1 GC2 GC3 GC Total GC2 0.0293 GC3 0.2164 −0.1271 GC Total 0.5335*** 0.3577*** 0.8178*** ENC −0.2017 0.0601 −0.8904*** −0.7549*** CN 0.0242 −0.0805 −0.1001 −0.1078 注:***表示P<0.001;GC1、GC2、GC3分别表示基因中所有密码子的第1位、第2位、第3位的GC含量;GC Total 表示密码子总GC含量;ENC表示有效密码子数;CN:密码子数。 2.5 PR2-plot分析
为了研究偏向密码子选择是否局限于高度偏向的蛋白质编码基因,通过PR2-plot分析了64个密码子氨基酸家族中嘌呤和嘧啶之间的关联[29],若密码子使用模式完全由突变造成,则G和C以及A和T的使用频率应相等。然而图5显示,在里氏木霉中G和C的使用频率高于A和T,说明里氏木霉密码子的使用模式除了核苷酸的组成,还受到其它因素的影响,例如选择压力等。
2.6 相对同义密码子使用分析
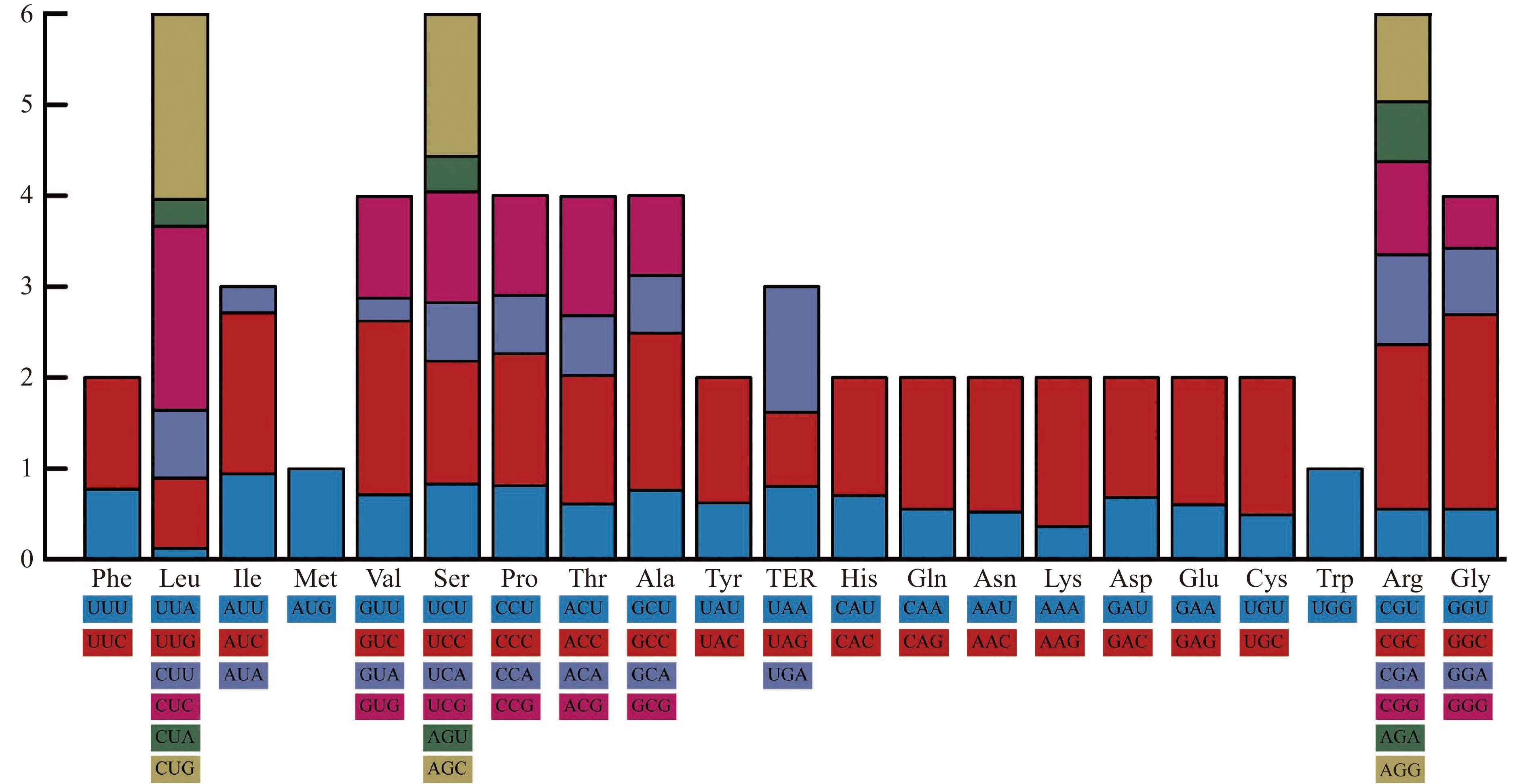
为了确定同义密码子的使用模式以及C/G末端密码子的首选程度,本文进行了相对同义密码子使用(RSCU)分析并计算了RSCU值(表2),绘制RSCU堆积图(图6)。在24个最常用的密码子中,22个(UUC、CUG、AUC、AUG、GUC、UCC、CCC、ACC、GCC、UAG、CAC、CAG、AAC、AAG、GAC、GAG、UCG、UGG、CGC、AGC、AGG、GGC)是C/G末端密码子(C末端:13个;G末端:9个),其余2个(UAA、AGA)是A末端密码子;没有一个首选密码子是U末端的。这些结果表明,核苷酸组成在里氏木霉密码子使用模式中起着不可或缺的作用。
表 2 里氏木霉蛋白编码区相对同义密码子使用度Table 2. RSCU analysis of protein coding region acid in T. reesei coding sequences氨基酸 密码子 数目 RSCU 氨基酸 密码子 数目 RSCU Phe UUU 65798 0.77 Tyr UAU 39376 0.62 UUC 105228 1.23 UAA 86733 1.38 Leu UUA 8336 0.12 TER UAA 2434 0.80 UUG 54088 0.77 UAG 2514 0.82 CUU 52375 0.75 His CAU 39404 0.70 CUC 141837 2.02 CAC 73860 1.30 CUA 20852 0.30 Gln CAA 52849 0.55 CUG 143771 2.05 CAG 138662 1.45 Ile AUU 69292 0.94 Asn AAU 42059 0.52 AUC 130569 1.77 AAC 121050 1.48 AUA 21446 0.29 Lys AAA 41032 0.36 Met AUG 103232 1.00 AAG 183878 1.64 Val GUU 51310 0.71 Asp GAU 93342 0.68 GUC 137774 1.91 GAC 180585 1.32 GUA 18067 0.25 Glu GAA 86554 0.60 GUG 80828 1.12 GAG 203315 1.40 Ser UCU 53294 0.83 Cys UGU 14149 0.49 UCC 86641 1.35 UGC 43486 1.51 UCA 41082 0.64 TER UGA 4232 1.38 UGG 78192 1.22 Trp UGG 68452 1.00 Pro CCU 58318 0.81 Arg CGU 26927 0.55 CCC 103910 1.45 CGC 88930 1.81 CCA 45925 0.64 CGA 48497 0.99 CCG 79130 1.10 CGG 50189 1.02 Thr ACU 41210 0.61 Ser AGU 24809 0.39 ACC 94630 1.41 AGC 100916 1.57 ACA 44135 0.66 Arg AGA 32260 0.66 ACG 88131 1.31 AGG 47231 0.96 Ala GCU 82619 0.76 Gly GGU 44800 0.55 GCC 187406 1.73 GGC 173318 2.14 GCA 68205 0.63 GGA 59183 0.73 GCG 95792 0.88 GGG 46294 0.57 注:大于等于1的RSCU值加粗表示。 2.7 高表达密码子的确定
以ENC值为偏好性标准,对基因进行排序,两极各取10%构建高低表达基因库,计算高低基因表达库密码子RSCU值和△RSCU值(表3),星号标注的21个密码子是高表达优越密码子,这些密码子(除了终止密码子UAA)全部以C或G结尾,这表明里氏木霉中的密码子使用偏向于C或G结尾的同义密码子。此外,4个密码子CUC、GCC、CGC和GGC是里氏木霉高表达基因的最优密码子。
表 3 里氏木霉最优密码子分析Table 3. Optimal codons in T. reeseiAA 密码子 H gene L gene △RSCU 氨基酸 密码子 H gene L gene △RSCU 数目 RSCU 数目 RSCU 数目 RSCU 数目 RSCU Phe UUU 3524 0.53 7108 0.89 −0.36 Tyr UAU 958 0.20 5522 0.94 −0.74 UUC* 9737 1.47 8883 1.11 0.36 UAC** 8835 1.80 6252 1.06 0.74 Leu UUA 75 0.02 2329 0.33 −0.31 TER UAA** 449 1.46 193 0.63 0.83 UUG 1166 0.24 7593 1.09 −0.85 UAG 176 0.57 310 1.01 −0.44 CUU 1241 0.25 8020 1.15 −0.90 His CAU 718 0.19 6238 1.03 −0.84 CUC*** 14466 2.96 9918 1.42 1.54 CAC** 6842 1.81 5819 0.97 0.84 CUA 256 0.05 4161 0.60 −0.55 Gln CAA 1082 0.19 8116 0.89 −0.70 CUG** 12109 2.48 9749 1.40 1.08 CAG** 10546 1.81 10083 1.11 0.70 Ile AUU 3628 0.65 7445 1.02 −0.37 Asn AAU 876 0.15 6230 0.86 −0.71 AUC** 12703 2.29 9975 1.37 0.92 AAC** 11157 1.85 8278 1.14 0.71 AUA 307 0.06 4449 0.61 −0.55 Lys AAA 855 0.10 6821 0.74 −0.64 Met AUG 7219 1.00 9316 1.00 0.00 AAG** 16815 1.90 11721 1.26 0.64 Val GUU 2093 0.36 6594 1.03 −0.67 Asp GAU 2606 0.28 11788 0.95 −0.67 GUC** 16087 2.80 8932 1.40 1.40 GAC** 15982 1.72 13077 1.05 0.67 GUA 250 0.04 3374 0.53 −0.49 Glu GAA 2057 0.21 11364 0.87 −0.66 GUG 4567 0.79 6666 1.04 −0.25 GAG** 17144 1.79 14830 1.13 0.66 Ser UCU 1997 0.50 6686 1.07 −0.57 Cys UGU 204 0.11 2923 0.79 −0.68 UCC** 8553 2.14 6236 1.00 1.14 UGC** 3613 1.89 4522 1.21 0.68 UCA 1042 0.26 6179 0.99 −0.73 TER UGA 295 0.96 415 1.36 −0.40 UCG 4870 1.22 6284 1.01 0.21 Trp UGG 4676 1.00 7218 1.00 0.00 Pro CCU 2256 0.49 6510 1.02 −0.53 Arg CGU 1332 0.44 3380 0.71 −0.27 CCC** 10969 2.40 5986 0.94 1.46 CGC*** 10626 3.51 5323 1.11 2.40 CCA 682 0.15 6996 1.10 −0.95 CGA 1347 0.45 5491 1.15 −0.70 CCG 4383 0.96 6060 0.95 0.01 CGG 2474 0.82 4373 0.92 −0.10 Thr ACU 1580 0.33 5457 0.88 −0.55 Ser AGU 375 0.09 4155 0.67 −0.58 ACC** 10733 2.21 6408 1.03 1.18 AGC** 7152 1.79 7940 1.27 0.52 ACA 1086 0.22 6555 1.06 −0.84 Arg AGA 521 0.17 5215 1.09 −0.92 ACG 6020 1.24 6373 1.03 0.21 AGG 1857 0.61 4888 1.02 −0.41 Ala GCU 3879 0.47 9349 1.00 −0.53 Gly GGU 2787 0.44 5157 0.75 −0.31 GCC*** 22460 2.69 10879 1.16 1.53 GGC*** 19097 3.04 10070 1.46 1.58 GCA 1488 0.18 9613 1.03 −0.85 GGA 1550 0.25 7121 1.03 −0.78 GCG 5518 0.66 7523 0.81 −0.15 GGG 1655 0.26 5264 0.76 −0.50 注:*表示△RSCU>0.3;**表示△RSCU>0.5;***表示△RSCU >1.5。 2.8 与其他真菌密码子偏好性比较
将里氏木霉分别与同属的长梗木霉、丝状模式真菌粗糙脉孢霉、模式真菌酿酒酵母的密码子使用频率进行比较(表4),其中R/L、R/N、R/S分别表示里氏木霉与长梗木霉、粗糙脉孢霉、酿酒酵母的每种密码子使用频率比值。结果显示,里氏木霉与酿酒酵母的密码子使用频率比值有34种大于等于2.0或小于等于0.5,占53.1%;与长梗木霉的密码子使用频率比值几乎都接近于1;而里氏木霉与粗糙脉孢霉的密码子使用频率比值有6种大于等于1.5或小于等于0.67,占9.3%。说明丝状真菌里氏木霉与模式真菌酿酒酵母的密码子偏好性差别较大,而与同属的长梗木霉以及丝状模式真菌粗糙脉孢霉的密码子偏好性差别相对较小。粗糙脉孢霉经常作为研究木质纤维素降解的模式真菌,将相关基因表达于里氏木霉时,两者密码子使用模式接近,无需考虑两者密码子的偏好性。
表 4 里氏木霉与其他物种密码子偏好性比较Table 4. Comparision of codon preference between T. reesei and other species氨基酸 密码子 密码子使用频率 频率比值 R L N S R/L R/N R/S Phe UUU 13.96 13.38 12.27 26.97 1.04 1.14 0.52 UUC 22.32 23.03 21.43 18.45 0.97 1.04 1.21 Leu UUA 1.77 1.74 2.83 26.33 1.02 0.63 0.07 UUG 11.47 11.64 15.42 27.00 0.99 0.74 0.42 CUU 11.11 11.53 14.34 12.63 0.96 0.77 0.88 CUC 30.08 30.39 26.23 5.73 0.99 1.15 5.25 CUA 4.42 4.39 6.05 13.50 1.01 0.73 0.33 CUG 30.49 30.52 18.31 10.87 1.00 1.67 2.80 Ile AUU 14.70 14.19 13.91 30.16 1.04 1.06 0.49 AUC 27.69 28.15 26.01 16.84 0.98 1.06 1.64 AUA 4.55 4.51 4.23 18.36 1.01 1.08 0.25 Met AUG 21.90 22.05 21.46 20.78 0.99 1.02 1.05 Val GUU 10.88 10.91 14.03 21.63 1.00 0.78 0.50 GUC 29.22 29.39 24.13 11.29 0.99 1.21 2.59 GUA 3.83 3.88 5.54 12.18 0.99 0.69 0.31 GUG 17.14 17.23 15.80 11.02 0.99 1.08 1.55 Ser UCU 11.30 11.04 12.24 23.49 1.02 0.92 0.48 UCC 18.38 18.47 20.01 14.18 1.00 0.92 1.30 UCA 8.71 8.65 9.57 18.89 1.01 0.91 0.46 UCG 16.58 16.66 14.94 8.86 1.00 1.11 1.87 Pro CCU 12.37 12.26 15.60 13.33 1.01 0.79 0.93 CCC 22.04 21.69 22.34 6.94 1.02 0.99 3.17 CCA 9.74 9.46 12.82 17.52 1.03 0.76 0.56 CCG 16.78 17.18 15.11 5.32 0.98 1.11 3.15 Thr ACU 8.74 8.49 11.41 19.98 1.03 0.77 0.44 ACC 20.07 19.92 24.74 12.40 1.01 0.81 1.62 ACA 9.36 8.92 11.14 17.75 1.05 0.84 0.53 ACG 18.69 18.84 13.92 8.23 0.99 1.34 2.27 Ala GCU 17.52 17.40 20.92 20.14 1.01 0.84 0.87 GCC 39.75 39.19 35.26 12.25 1.01 1.13 3.24 GCA 14.47 14.08 13.00 16.26 1.03 1.11 0.89 GCG 20.32 21.15 17.47 6.34 0.96 1.16 3.21 Tyr UAU 8.35 8.30 8.62 18.90 1.01 0.97 0.44 UAC 18.40 18.30 16.94 14.39 1.01 1.09 1.28 TER UUA 0.52 0.42 0.54 1.00 1.24 0.96 0.52 UAG 0.53 0.47 0.56 0.49 1.13 0.95 1.09 His CAU 8.36 8.47 9.84 13.68 0.99 0.85 0.61 CAC 15.67 15.63 14.75 7.61 1.00 1.06 2.06 Gln CAA 11.21 11.14 17.29 26.50 1.01 0.65 0.42 CAG 29.41 29.36 25.47 12.34 1.00 1.15 2.38 Asn AAU 8.92 8.78 10.69 36.10 1.02 0.83 0.25 AAC 25.67 25.54 26.34 24.40 1.01 0.97 1.05 Lys AAA 8.70 8.79 11.94 42.22 0.99 0.73 0.21 AAG 39.00 38.72 38.66 30.42 1.01 1.01 1.28 Asp GAU 19.80 19.98 24.31 37.77 0.99 0.81 0.52 GAC 38.30 38.20 32.21 20.01 1.00 1.19 1.91 Glu GAA 18.36 18.45 23.16 45.43 1.00 0.79 0.40 GAG 43.12 43.18 41.78 19.59 1.00 1.03 2.20 Cys UGU 3.00 3.06 3.52 8.18 0.98 0.85 0.37 UGC 9.22 9.32 7.45 5.00 0.99 1.24 1.85 TER UGA 0.90 0.83 0.76 0.63 1.08 1.18 1.42 Trp UGG 14.52 14.45 13.31 10.49 1.00 1.09 1.38 Arg CGU 5.71 5.87 8.57 6.26 0.97 0.67 0.91 CGC 18.86 18.53 17.34 2.70 1.02 1.09 7.00 CGA 10.29 10.56 7.18 3.14 0.97 1.43 3.28 CGG 10.65 11.13 8.52 1.91 0.96 1.25 5.58 Ser AGU 5.26 5.21 8.90 14.63 1.01 0.59 0.36 AGC 21.40 21.16 17.79 10.17 1.01 1.20 2.10 Arg AGA 6.84 6.81 8.00 20.94 1.00 0.86 0.33 AGG 10.02 10.26 11.83 9.62 0.98 0.85 1.04 Gly GGU 9.50 9.83 17.50 22.50 0.97 0.54 0.42 GGC 36.76 36.08 28.47 9.90 1.02 1.29 3.71 GGA 12.55 12.68 13.82 11.24 0.99 0.91 1.12 GGG 9.82 10.18 11.45 6.23 0.96 0.86 1.58 注:与粗糙脉孢霉密码子使用频率比值≥1.50 或≤0.67的用下划线标记;与酿酒酵母密码子使用频率比值≥2.00 或≤0.5的用粗体标记;字母R、L、N、S分别表示里氏木霉、长梗木霉、粗糙脉孢霉、酿酒酵母。 3. 讨论与结论
当重组蛋白异源表达时,密码子使用偏好对蛋白质表达水平有重要的影响。DNA序列中密码子的频率与物种中相应的tRNA呈正相关,tRNA浓度决定了可用于蛋白质翻译延伸的氨基酸数量,进而影响蛋白质合成的效率[30]。蛋白质的表达水平与密码子使用偏好高度相关。稀有密码子往往会降低翻译速度,甚至导致翻译错误。因此,密码子优化是增加蛋白质表达的最关键的决定因素。
里氏木霉作为工业生产纤维素酶的菌株,其某些突变株的蛋白分泌能力在发酵条件下可达到100 g/L[31-32],鉴于此优良特征,里氏木霉可以作为异源蛋白表达的优良宿主。对其密码子偏好性进行研究具有重要的理论研究和工业应用意义。在本研究中,通过对里氏木霉基因组进行分析,编码区的GC3(70.4%)含量表明,该基因组富含C+G,总体密码子使用偏向于C和G末端密码子。在进化过程中,若A(T)到G(C)的突变压力大,那么密码子的第3位碱基是G(C)的概率就要高[33]。在里氏木霉使用频率较高的24个密码子中,有22个均是以GC结尾的。通过对里氏木霉基因组密码子使用模式的分析发现,其密码子使用的偏好性受到选择压力的影响,其次自然选择在塑造密码子偏好性使用过程中也扮演着非常重要的作用。通过ENC差异构建了里氏木霉高低表达基因库,确定了21个高表达优越密码子和4个高表达最优密码子(CUC、GCC、CGC和GGC)。
将里氏木霉分别与其它真菌的密码子使用频率进行比较,发现里氏木霉基因的密码子偏好性与酵母的差异较大,这可以解释为什么里氏木霉的许多基因都无法实现在毕赤酵母中的异源表达,然而通过对来源于里氏木霉的Cel5A、Cel6A经过密码子优化后,可以成功在毕赤酵母中进行表达[34,35]。里氏木霉与粗糙脉孢霉的密码子偏好性差异最小,因此不经过任何密码子优化的里氏木霉基因可以在粗糙脉孢霉中成功表达并且互补粗糙脉孢霉相关基因的缺失表型[36]。这些例子充分表明密码子偏好性对基因表达的重要性。本研究对里氏木霉的密码子使用偏好性进行了系统分析,可为外源基因在里氏木霉以及与其进化关系较为接近的其他物种中进行异源表达时提供密码子优化指导。
-
![]()
图 4 有效密码子数(ENC)比率的频率分布图
Figure 4. Frequency distribution of the effective number of codons (ENC) ratio
表 1 各基因相关参数的相关性分析
Table 1 Correlation analysis of each gene-related parameters
指标 GC1 GC2 GC3 GC Total GC2 0.0293 GC3 0.2164 −0.1271 GC Total 0.5335*** 0.3577*** 0.8178*** ENC −0.2017 0.0601 −0.8904*** −0.7549*** CN 0.0242 −0.0805 −0.1001 −0.1078 注:***表示P<0.001;GC1、GC2、GC3分别表示基因中所有密码子的第1位、第2位、第3位的GC含量;GC Total 表示密码子总GC含量;ENC表示有效密码子数;CN:密码子数。  下载: 导出CSV
下载: 导出CSV
表 2 里氏木霉蛋白编码区相对同义密码子使用度
Table 2 RSCU analysis of protein coding region acid in T. reesei coding sequences
氨基酸 密码子 数目 RSCU 氨基酸 密码子 数目 RSCU Phe UUU 65798 0.77 Tyr UAU 39376 0.62 UUC 105228 1.23 UAA 86733 1.38 Leu UUA 8336 0.12 TER UAA 2434 0.80 UUG 54088 0.77 UAG 2514 0.82 CUU 52375 0.75 His CAU 39404 0.70 CUC 141837 2.02 CAC 73860 1.30 CUA 20852 0.30 Gln CAA 52849 0.55 CUG 143771 2.05 CAG 138662 1.45 Ile AUU 69292 0.94 Asn AAU 42059 0.52 AUC 130569 1.77 AAC 121050 1.48 AUA 21446 0.29 Lys AAA 41032 0.36 Met AUG 103232 1.00 AAG 183878 1.64 Val GUU 51310 0.71 Asp GAU 93342 0.68 GUC 137774 1.91 GAC 180585 1.32 GUA 18067 0.25 Glu GAA 86554 0.60 GUG 80828 1.12 GAG 203315 1.40 Ser UCU 53294 0.83 Cys UGU 14149 0.49 UCC 86641 1.35 UGC 43486 1.51 UCA 41082 0.64 TER UGA 4232 1.38 UGG 78192 1.22 Trp UGG 68452 1.00 Pro CCU 58318 0.81 Arg CGU 26927 0.55 CCC 103910 1.45 CGC 88930 1.81 CCA 45925 0.64 CGA 48497 0.99 CCG 79130 1.10 CGG 50189 1.02 Thr ACU 41210 0.61 Ser AGU 24809 0.39 ACC 94630 1.41 AGC 100916 1.57 ACA 44135 0.66 Arg AGA 32260 0.66 ACG 88131 1.31 AGG 47231 0.96 Ala GCU 82619 0.76 Gly GGU 44800 0.55 GCC 187406 1.73 GGC 173318 2.14 GCA 68205 0.63 GGA 59183 0.73 GCG 95792 0.88 GGG 46294 0.57 注:大于等于1的RSCU值加粗表示。
下载: 导出CSV
表 3 里氏木霉最优密码子分析
Table 3 Optimal codons in T. reesei
AA 密码子 H gene L gene △RSCU 氨基酸 密码子 H gene L gene △RSCU 数目 RSCU 数目 RSCU 数目 RSCU 数目 RSCU Phe UUU 3524 0.53 7108 0.89 −0.36 Tyr UAU 958 0.20 5522 0.94 −0.74 UUC* 9737 1.47 8883 1.11 0.36 UAC** 8835 1.80 6252 1.06 0.74 Leu UUA 75 0.02 2329 0.33 −0.31 TER UAA** 449 1.46 193 0.63 0.83 UUG 1166 0.24 7593 1.09 −0.85 UAG 176 0.57 310 1.01 −0.44 CUU 1241 0.25 8020 1.15 −0.90 His CAU 718 0.19 6238 1.03 −0.84 CUC*** 14466 2.96 9918 1.42 1.54 CAC** 6842 1.81 5819 0.97 0.84 CUA 256 0.05 4161 0.60 −0.55 Gln CAA 1082 0.19 8116 0.89 −0.70 CUG** 12109 2.48 9749 1.40 1.08 CAG** 10546 1.81 10083 1.11 0.70 Ile AUU 3628 0.65 7445 1.02 −0.37 Asn AAU 876 0.15 6230 0.86 −0.71 AUC** 12703 2.29 9975 1.37 0.92 AAC** 11157 1.85 8278 1.14 0.71 AUA 307 0.06 4449 0.61 −0.55 Lys AAA 855 0.10 6821 0.74 −0.64 Met AUG 7219 1.00 9316 1.00 0.00 AAG** 16815 1.90 11721 1.26 0.64 Val GUU 2093 0.36 6594 1.03 −0.67 Asp GAU 2606 0.28 11788 0.95 −0.67 GUC** 16087 2.80 8932 1.40 1.40 GAC** 15982 1.72 13077 1.05 0.67 GUA 250 0.04 3374 0.53 −0.49 Glu GAA 2057 0.21 11364 0.87 −0.66 GUG 4567 0.79 6666 1.04 −0.25 GAG** 17144 1.79 14830 1.13 0.66 Ser UCU 1997 0.50 6686 1.07 −0.57 Cys UGU 204 0.11 2923 0.79 −0.68 UCC** 8553 2.14 6236 1.00 1.14 UGC** 3613 1.89 4522 1.21 0.68 UCA 1042 0.26 6179 0.99 −0.73 TER UGA 295 0.96 415 1.36 −0.40 UCG 4870 1.22 6284 1.01 0.21 Trp UGG 4676 1.00 7218 1.00 0.00 Pro CCU 2256 0.49 6510 1.02 −0.53 Arg CGU 1332 0.44 3380 0.71 −0.27 CCC** 10969 2.40 5986 0.94 1.46 CGC*** 10626 3.51 5323 1.11 2.40 CCA 682 0.15 6996 1.10 −0.95 CGA 1347 0.45 5491 1.15 −0.70 CCG 4383 0.96 6060 0.95 0.01 CGG 2474 0.82 4373 0.92 −0.10 Thr ACU 1580 0.33 5457 0.88 −0.55 Ser AGU 375 0.09 4155 0.67 −0.58 ACC** 10733 2.21 6408 1.03 1.18 AGC** 7152 1.79 7940 1.27 0.52 ACA 1086 0.22 6555 1.06 −0.84 Arg AGA 521 0.17 5215 1.09 −0.92 ACG 6020 1.24 6373 1.03 0.21 AGG 1857 0.61 4888 1.02 −0.41 Ala GCU 3879 0.47 9349 1.00 −0.53 Gly GGU 2787 0.44 5157 0.75 −0.31 GCC*** 22460 2.69 10879 1.16 1.53 GGC*** 19097 3.04 10070 1.46 1.58 GCA 1488 0.18 9613 1.03 −0.85 GGA 1550 0.25 7121 1.03 −0.78 GCG 5518 0.66 7523 0.81 −0.15 GGG 1655 0.26 5264 0.76 −0.50 注:*表示△RSCU>0.3;**表示△RSCU>0.5;***表示△RSCU >1.5。
下载: 导出CSV
表 4 里氏木霉与其他物种密码子偏好性比较
Table 4 Comparision of codon preference between T. reesei and other species
氨基酸 密码子 密码子使用频率 频率比值 R L N S R/L R/N R/S Phe UUU 13.96 13.38 12.27 26.97 1.04 1.14 0.52 UUC 22.32 23.03 21.43 18.45 0.97 1.04 1.21 Leu UUA 1.77 1.74 2.83 26.33 1.02 0.63 0.07 UUG 11.47 11.64 15.42 27.00 0.99 0.74 0.42 CUU 11.11 11.53 14.34 12.63 0.96 0.77 0.88 CUC 30.08 30.39 26.23 5.73 0.99 1.15 5.25 CUA 4.42 4.39 6.05 13.50 1.01 0.73 0.33 CUG 30.49 30.52 18.31 10.87 1.00 1.67 2.80 Ile AUU 14.70 14.19 13.91 30.16 1.04 1.06 0.49 AUC 27.69 28.15 26.01 16.84 0.98 1.06 1.64 AUA 4.55 4.51 4.23 18.36 1.01 1.08 0.25 Met AUG 21.90 22.05 21.46 20.78 0.99 1.02 1.05 Val GUU 10.88 10.91 14.03 21.63 1.00 0.78 0.50 GUC 29.22 29.39 24.13 11.29 0.99 1.21 2.59 GUA 3.83 3.88 5.54 12.18 0.99 0.69 0.31 GUG 17.14 17.23 15.80 11.02 0.99 1.08 1.55 Ser UCU 11.30 11.04 12.24 23.49 1.02 0.92 0.48 UCC 18.38 18.47 20.01 14.18 1.00 0.92 1.30 UCA 8.71 8.65 9.57 18.89 1.01 0.91 0.46 UCG 16.58 16.66 14.94 8.86 1.00 1.11 1.87 Pro CCU 12.37 12.26 15.60 13.33 1.01 0.79 0.93 CCC 22.04 21.69 22.34 6.94 1.02 0.99 3.17 CCA 9.74 9.46 12.82 17.52 1.03 0.76 0.56 CCG 16.78 17.18 15.11 5.32 0.98 1.11 3.15 Thr ACU 8.74 8.49 11.41 19.98 1.03 0.77 0.44 ACC 20.07 19.92 24.74 12.40 1.01 0.81 1.62 ACA 9.36 8.92 11.14 17.75 1.05 0.84 0.53 ACG 18.69 18.84 13.92 8.23 0.99 1.34 2.27 Ala GCU 17.52 17.40 20.92 20.14 1.01 0.84 0.87 GCC 39.75 39.19 35.26 12.25 1.01 1.13 3.24 GCA 14.47 14.08 13.00 16.26 1.03 1.11 0.89 GCG 20.32 21.15 17.47 6.34 0.96 1.16 3.21 Tyr UAU 8.35 8.30 8.62 18.90 1.01 0.97 0.44 UAC 18.40 18.30 16.94 14.39 1.01 1.09 1.28 TER UUA 0.52 0.42 0.54 1.00 1.24 0.96 0.52 UAG 0.53 0.47 0.56 0.49 1.13 0.95 1.09 His CAU 8.36 8.47 9.84 13.68 0.99 0.85 0.61 CAC 15.67 15.63 14.75 7.61 1.00 1.06 2.06 Gln CAA 11.21 11.14 17.29 26.50 1.01 0.65 0.42 CAG 29.41 29.36 25.47 12.34 1.00 1.15 2.38 Asn AAU 8.92 8.78 10.69 36.10 1.02 0.83 0.25 AAC 25.67 25.54 26.34 24.40 1.01 0.97 1.05 Lys AAA 8.70 8.79 11.94 42.22 0.99 0.73 0.21 AAG 39.00 38.72 38.66 30.42 1.01 1.01 1.28 Asp GAU 19.80 19.98 24.31 37.77 0.99 0.81 0.52 GAC 38.30 38.20 32.21 20.01 1.00 1.19 1.91 Glu GAA 18.36 18.45 23.16 45.43 1.00 0.79 0.40 GAG 43.12 43.18 41.78 19.59 1.00 1.03 2.20 Cys UGU 3.00 3.06 3.52 8.18 0.98 0.85 0.37 UGC 9.22 9.32 7.45 5.00 0.99 1.24 1.85 TER UGA 0.90 0.83 0.76 0.63 1.08 1.18 1.42 Trp UGG 14.52 14.45 13.31 10.49 1.00 1.09 1.38 Arg CGU 5.71 5.87 8.57 6.26 0.97 0.67 0.91 CGC 18.86 18.53 17.34 2.70 1.02 1.09 7.00 CGA 10.29 10.56 7.18 3.14 0.97 1.43 3.28 CGG 10.65 11.13 8.52 1.91 0.96 1.25 5.58 Ser AGU 5.26 5.21 8.90 14.63 1.01 0.59 0.36 AGC 21.40 21.16 17.79 10.17 1.01 1.20 2.10 Arg AGA 6.84 6.81 8.00 20.94 1.00 0.86 0.33 AGG 10.02 10.26 11.83 9.62 0.98 0.85 1.04 Gly GGU 9.50 9.83 17.50 22.50 0.97 0.54 0.42 GGC 36.76 36.08 28.47 9.90 1.02 1.29 3.71 GGA 12.55 12.68 13.82 11.24 0.99 0.91 1.12 GGG 9.82 10.18 11.45 6.23 0.96 0.86 1.58 注:与粗糙脉孢霉密码子使用频率比值≥1.50 或≤0.67的用下划线标记;与酿酒酵母密码子使用频率比值≥2.00 或≤0.5的用粗体标记;字母R、L、N、S分别表示里氏木霉、长梗木霉、粗糙脉孢霉、酿酒酵母。
下载: 导出CSV
-
[1] KEMURA T. Codon usage and tRNA content in unicellular and multicellular organisms[J]. Molecular Biology and Evolution,1985,2:13−34.
[2] DAS S, ROYMONDAL U, SAHOO S. Analyzing gene expression from relative codon usage bias in yeast genome: A statistical significance and biological relevance[J]. Gene,2009,443(1-2):121−31. doi: 10.1016/j.gene.2009.04.022
[3] MORIYAMA E N. Codon usage bias and tRNA Abundance in Drosophila[J]. Journal of Molecular Evolution,1997,45(5):514−523. doi: 10.1007/PL00006256
[4] DURET L, MOUCHIROUD D. Expression pattern and, surprisingly, gene length shape codon usage in Caenorhabditis, Drosophila, and Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(8):4482−4487. doi: 10.1073/pnas.96.8.4482
[5] MORIYAMA E. Gene length and codon usage bias in Drosophila melanogaster, Saccharomyces cerevisiae and Escherichia coli[J]. Nucleic Acids Research,1998,26(19):3188.
[6] GUPTA S K, MAJUMDAR S, BHATTACHARYA T K, et al. Studies on the relationships between the synonymous codon usage and protein secondary structural units[J]. Biochemical and Biophysical Research Communications,2000,269(3):692−696. doi: 10.1006/bbrc.2000.2351
[7] SHARP P M, BAILES E, GROCOCK R J, et al. Variation in the strength of selected codon usage bias among bacteria[J]. Nucleic Acids Resarch,2005,33(4):1141−1153. doi: 10.1093/nar/gki242
[8] SHARP P M, TUOHY T M, MOSURSKI K R. Codon usage in yeast: Cluster analysis clearly differentiates highly and lowly expressed genes[J]. Nucleic Acids Resarch,1986,14(13):5125−5143. doi: 10.1093/nar/14.13.5125
[9] ZHOU T, SUN X, LU Z. Synonymous codon usage in environmental chlamydia UWE25 reflects an evolutional divergence from pathogenic chlamydiae[J]. Gene,2006,368:117−125. doi: 10.1016/j.gene.2005.10.035
[10] CHIAPELLO H, LISACEK F, CABOCHE M, et al. Codon usage and gene function are related in sequences of Arabidopsis thaliana[J]. Gene,1998,209(1):1−38.
[11] SHARP P M, ELIZABETH C, HIGGINS D G, et al. Codon usage patterns in Escherichia coli, Bacillus subtilis, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Drosophila melanogaster and Homo sapiens; a review of the considerable within-species diversity[J]. Nucleic Acids Resarch,1988,16(17):8207−8211. doi: 10.1093/nar/16.17.8207
[12] BISCHOF R H, RAMONI J, SEIBOTH B. Cellulases and beyond: the first 70 years of the enzyme producer Trichoderma reesei[J]. Microbial Cell Factories,2016,15(1):106. doi: 10.1186/s12934-016-0507-6
[13] ZIELIŃSKA D, SZENTNER K, WAśKIEWICZ A, et al. Production of nanocellulose by enzymatic treatment for application in polymer composites[J]. Materials,2021,14(9):2124. doi: 10.3390/ma14092124
[14] KUMAR M R, KUMARAN M D, BALASHANMUGAM P. Production of cellulase enzyme by Trichoderma reesei Cefl9 and its application in the production of bio-ethanol[J]. Pakistan Journal of Pharmaceutical Sciences,2014,17(5):735−739.
[15] PEIL S, BECKERS S J, FISCHER J, et al. Biodegradable, lignin-based encapsulation enables delivery of Trichoderma reesei with programmed enzymatic release against grapevine trunk diseases[J]. Materials Today Bio,2020,7:100061. doi: 10.1016/j.mtbio.2020.100061
[16] LICHTENBERG J, PEREZ C E, MADSEN K, et al. Safety evaluation of a novel muramidase for feed application[J]. Regulatory Toxicology and Pharmacology,2017,89:57−69. doi: 10.1016/j.yrtph.2017.07.014
[17] LANDOWSKI C P, HUUSKONEN A, WAHL R, et al. Enabling low cost biopharmaceuticals: a systematic approach to delete proteases from a well-known protein production host Trichoderma reesei[J]. PLoS One,2015,10(8):e0134723. doi: 10.1371/journal.pone.0134723
[18] SALOHEIMO M, CULLEN D, MARTINEZ D, et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina)[J]. Nature Biotechnology,2008,26(5):553−560. doi: 10.1038/nbt1403
[19] MICHAEL S. ROSENBERG M S, SUBRAMANIAN S, KUMAR S. Patterns of transitional mutation biases within and among mammalian genomes[J]. Molecular Biology and Evolution,2003,20(6):988−993. doi: 10.1093/molbev/msg113
[20] SUEOKA N. Directional mutation pressure and neutral molecular evolution[J]. Proceedings of the National Academy of Sciences,1988,85:2653−2657. doi: 10.1073/pnas.85.8.2653
[21] WRIGHT F. The 'effective number of codons' used in a gene[J]. Gene,1990,87(1):23−29. doi: 10.1016/0378-1119(90)90491-9
[22] LIU H, RUI H, ZHANG H, et al. Analysis of synonymous codon usage in Zea mays[J]. Molecular Biology Reports,2010,37(2):677−684. doi: 10.1007/s11033-009-9521-7
[23] SUEOKA N. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses[J]. Journal of Molecular Evolution,2001,53(4-5):469−476. doi: 10.1007/s002390010237
[24] FUGLSANG A. The 'effective number of codons' revisited[J]. Biochemical and Biophysical Research Communications,2004,317(3):957−964. doi: 10.1016/j.bbrc.2004.03.138
[25] 刘庆坡, 薛庆中. 粳稻叶绿体基因组的密码子用法[J]. 作物学报, 2004, 30(12): 1220-1224. LIU Q P, XUE Q Z. Codon usage in the chloroplast genome of rice (Oryza sativa L. ssp. japonica)[J]. Acta Agronomica Sinica, 2004, 30(12); 1220-1224.
[26] ZHANG W J, JIE Z, LI Z F, et al. Comparative analysis of codon usage patterns among mitochondrion, chloroplast and nuclear Genes in Triticum aestivum L[J]. Journal of Integrative Plant Biology,2007,49(2):246−254. doi: 10.1111/j.1744-7909.2007.00404.x
[27] 范三红, 郭蔼光, 单丽伟, 等. 拟南芥基因密码子偏爱性分析[J]. 生物化学与生物物理进展,2003,30(2):221−225. [FAN S H, GUO A G, SHAN L W, et al. Analysis of genetic code preference in Arabidopsis thaliana[J]. Progress in Biochemistry and Biophysics,2003,30(2):221−225. doi: 10.3321/j.issn:1000-3282.2003.02.012 [28] JENKINS G M, HOLMES E C. The extent of codon usage bias in human RNA viruses and its evolutionary origin[J]. Virus Research,2003,92(1):1−7. doi: 10.1016/S0168-1702(02)00309-X
[29] SUEOKA N. Translation-coupled violation of Parity Rule 2 in human genes is not the cause of heterogeneity of the DNA G+C content of third codon position[J]. Gene,1999,238(1):53−58. doi: 10.1016/S0378-1119(99)00320-0
[30] GUSTAFSSON C, GOVINDARAJAN S, MINSHULL J. Codon bias and heterologous protein expression[J]. Trends is Biotechnology,2004,22(7):346−53. doi: 10.1016/j.tibtech.2004.04.006
[31] CHERRY J R, FIDANSTSEF A L. Directed evolution of industrial enzymes: an update[J]. Current Opinion in Biotechnology,2003,14(4):438−443. doi: 10.1016/S0958-1669(03)00099-5
[32] VISSER H, JOOSTEN V, PUNT P J, et al. Development of a mature fungal technology and production platform for industrial enzymes based on a Myceliophthora thermophila isolate, previously known as Chrysosporium lucknowense C1[J]. Industrial Biotechnology,2011,7(3):214−223. doi: 10.1089/ind.2011.7.214
[33] NOVEMBRE J A. Accounting for background nucleotide composition when measuring codon usage bias[J]. Molecular Biology and Evolution,2002(8):1390.
[34] SUN F F, BAI R, YANG H, et al. Heterologous expression of codon optimized Trichoderma reesei Cel6A in Pichia pastoris[J]. Enzyme and Microbial Technology,2016,92:107−116. doi: 10.1016/j.enzmictec.2016.07.004
[35] 白仁惠, 张云博, 王春迪, 等. 里氏木霉Cel5A基因优化及其在毕赤酵母中的高效表达[J]. 生物工程学报,2016,32(10):1381−1394. [BAI R H, ZHANG Y B, WANG C D, et al. Gene optimization and efficient expression of Trichoderma reesei Cel5A in Pichia pastoris[J]. Chinese Journal of Biotechnology,2016,32(10):1381−1394. [36] XIONG Y, WU V W, LUBBE A, et al. A fungal transcription factor essential for starch degradation affects integration of carbon and nitrogen metabolism[J]. PLoS Genetics,2017,13(5):e1006737. doi: 10.1371/journal.pgen.1006737

 下载:
下载:






计量
- 文章访问数: 301
- HTML全文浏览量: 235
- PDF下载量: 39
