


Construction of High-yield and High-purity Pectin Methylesterase Aspergillus niger Engineering Strain
-
摘要: 为获得一株高产高纯度的果胶甲酯酶的黑曲霉工程菌,提高果胶甲酯酶的产量,从果胶酶生产菌种中克隆了果胶甲酯酶基因pmeA,通过同源重组的原理,冻融法转化农杆菌、农杆菌介导法转化黑曲霉方法,成功构建了分泌表达果胶甲酯酶的纯合重组菌株TH-2(glaA::pmeA)。基本发酵培养基中发酵第9 d上清中最高酶活达到467.77 U/mL。进一步敲除重组菌株TH-2(glaA::pmeA)中背景蛋白酸稳定的α-淀粉酶的编码基因asaA,获得纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)。该菌株在添加1%的硫酸铵的发酵培养基中培养7 d后,发酵液上清中主要的背景蛋白均消失。但是与纯合重组菌株TH-2(glaA::pmeA)相比,果胶甲酯酶表达量有所下降,最高酶活为255.40 U/mL。重组果胶甲酯酶的最适作用温度为50 ℃,适合的温度范围是40~80 ℃,在80 ℃下仍能维持其酶活性的70%以上,适合的pH范围是3.0~5.0,最适pH为4.0。最终获得了一株温度和pH作用范围较宽的高产高纯度果胶甲酯酶的黑曲霉工程菌。Abstract: In order to obtain an engineering strain of Aspergillus niger with high yield and purity of pectin methylesterase and improve the yield of pectin methylesterase, the pectin methylesterase gene pmeA was cloned from the pectinase-producing strain, and Aspergillus niger was transformed by agrobacterium-mediated method, the homozygous recombinant strain TH-2 (glaA::pmeA). The highest enzyme activity in the supernatant on the 9th day of fermentation in the basic fermentation medium reached 467.77 U/mL. At the same time, SDS-PAGE showed that the main background protein α-amylase disappeared, but the acid-stable α-amylase remained. Further the background protein acid-stable α-amylase encoding gene asaA in the recombinant strain TH-2 (glaA::pmeA) was knock out to obtain the homozygous recombinant strain TH-2 (glaA::pmeA∆pyrG∆asaA). After the strain was cultured in a fermentation medium supplemented with 1% ammonium sulfate for 7 days, the main background proteins in the supernatant of the fermentation broth disappeared. However, compared with the homozygous recombinant strain TH-2 (glaA::pmeA), the expression of pectin methylesterase decreased, and the highest enzyme activity was 255.40 U/mL. The optimum temperature of the recombinant pectin methylesteraseis 50 ℃, the suitable temperature range was 40~80 ℃, it could still maintain more than 70% of its enzyme activity at 80 ℃, the suitable pH range was 3.0~5.0, the optimum pH was 4.0. Finally, an engineering strain of Aspergillus niger with high yield and high purity pectin methylesterase was obtained.
-
Keywords:
- pectin methylesterase /
- Aspergillus niger /
- background protein /
- secretory expression
-
果胶甲酯酶对果胶分子中的甲酯基团具有高度专一性,可以将其催化水解形成果胶酸,并释放氢离子和甲醇。由于具有这一特性,果胶甲酯酶在工业当中得到广泛的应用[1-2]。果胶甲酯酶可以将高酯果胶变成低酯果胶,低酯果胶是具有较高价值的工业原料[3],低酯果胶还可作为药物的组成成分,加快伤口愈合,减少血浆中的胆固醇。目前生产低酯果胶的最佳方法是果胶甲酯酶法生产,而低酯果胶产品的品质高度依赖于果胶甲酯酶制剂的纯度[4-6]。果胶甲酯酶还应用于造纸工业的白水处理[7-8],在饲料工业中它与其他酶系复合使用还可以提高饲料转化利用率[9]。此外由于果胶甲酯酶在罐头、果汁、泡菜等食品行业有较多的应用[10-12],因此,食品级果胶甲酯酶的需求量也在逐年增加,食品级果胶甲酯酶商品纯度的要求也很高[13-14]。
目前市面上销售的果胶甲酯酶多为黑曲霉发酵生产[15],而黑曲霉在发酵生产果胶甲酯酶的过程当中还同时分泌许多其他酶类,这就导致了在下游分离纯化果胶甲酯酶的过程当中产生产量的损失以及酶活的损耗[16-17]。同时分离纯化的工艺步骤也大大增加了产酶的成本,使得果胶甲酯酶的价格较高,并且目前果胶甲酯酶的分离纯化工艺也并不完善,最终的产品纯度也得不到保证[18],这对于一些高度依赖于果胶甲酯酶纯度的产品的生产是不利的。所以,获得高产高纯度果胶甲酯酶的工程菌具有重大的商业价值[19-20]。
本实验从果胶酶生产菌株黑曲霉种克隆了果胶甲酯酶基因pmeA,构建了由PglaA6R启动子和SglaA信号肽调控的表达载体pSZHG6R-pmeA,通过农杆菌介导法转化糖化酶生产菌黑曲霉TH-2,获得高效分泌表达果胶甲酯酶的纯合重组菌株TH-2(glaA::pmeA)。进一步通过敲除重组菌株TH-2(glaA::pmeA)中的asaA基因和优化发酵培养基,去除了背景蛋白,从而获得了高产高纯度果胶甲酯酶的工程菌。
1. 材料与方法
1.1 材料与仪器
黑曲霉TH-2、黑曲霉果胶酶生产菌、农杆菌AGL1、AGL1(pSZH-∆pyrG)、AGL1(pSZHA-pyrG),大肠杆菌DH5α、载体pSZHG6R 由真菌遗传研究室保存;限制性核酸内切酶、T4 DNA Ligase Takara公司;基因组DNA提取试剂盒、质粒提取试剂盒、琼脂糖凝胶回收试剂盒、SDS-PAGE凝胶制备试剂盒 康为世纪公司;利福平、卡那霉素、氨苄霉素、乙酰丁香酮、PCR引物(表1) 生工生物公司。
表 1 PCR引物序列Table 1. PCR Primer sequence引物名称 引物序列 P1:pmeA-sense TCTAGACGCATGACGGCTCCTT P2:pmeA-antisense AAGCTTAGATCTTAGTTGATGTAGCTAGTATC P3:5’GLA-sense TCTAGACTCGGCGACTTGGTCTTCAC P4:3’GLA-antisense CTCGAGCATAAGGCGGGTTCACATC P5:5’pyrG-sense GAATTCCTCGAGCTAACATACATTCCG P6:3’pyrG-antisense TCTAGAGGTAAGTAATCAGTAACTG P7:5’asaA-sence TCGCCTATCTACTCCTCATCC P8:3’asaA-antisence CTTCAACCCTGCTGCTAAATC PCR仪 杭州晶格科学仪器有限公司;T960型热循环仪、SDS-PAGE电泳系统 美国伯乐生命医学产品有限公司;Mini-PROTEAN® Tetra蛋白质电泳系统、凝胶成像系统 上海天能科技有限公司;Tanon 2500R全自动数码凝胶图像分析系统、控温水槽 上海博迅实业有限公司;DK-8D三温三控水槽、紫外分光光度计 上海元析仪器有限公司; UV-5800PC型紫外可见分光光度计、恒温振荡培养箱 美国精骐科技与产业有限公司;IS-RDD3型台式恒温振荡器、高压蒸汽灭菌锅 美国致微仪器有限公司。
1.2 实验方法
1.2.1 果胶甲酯酶基因的扩增
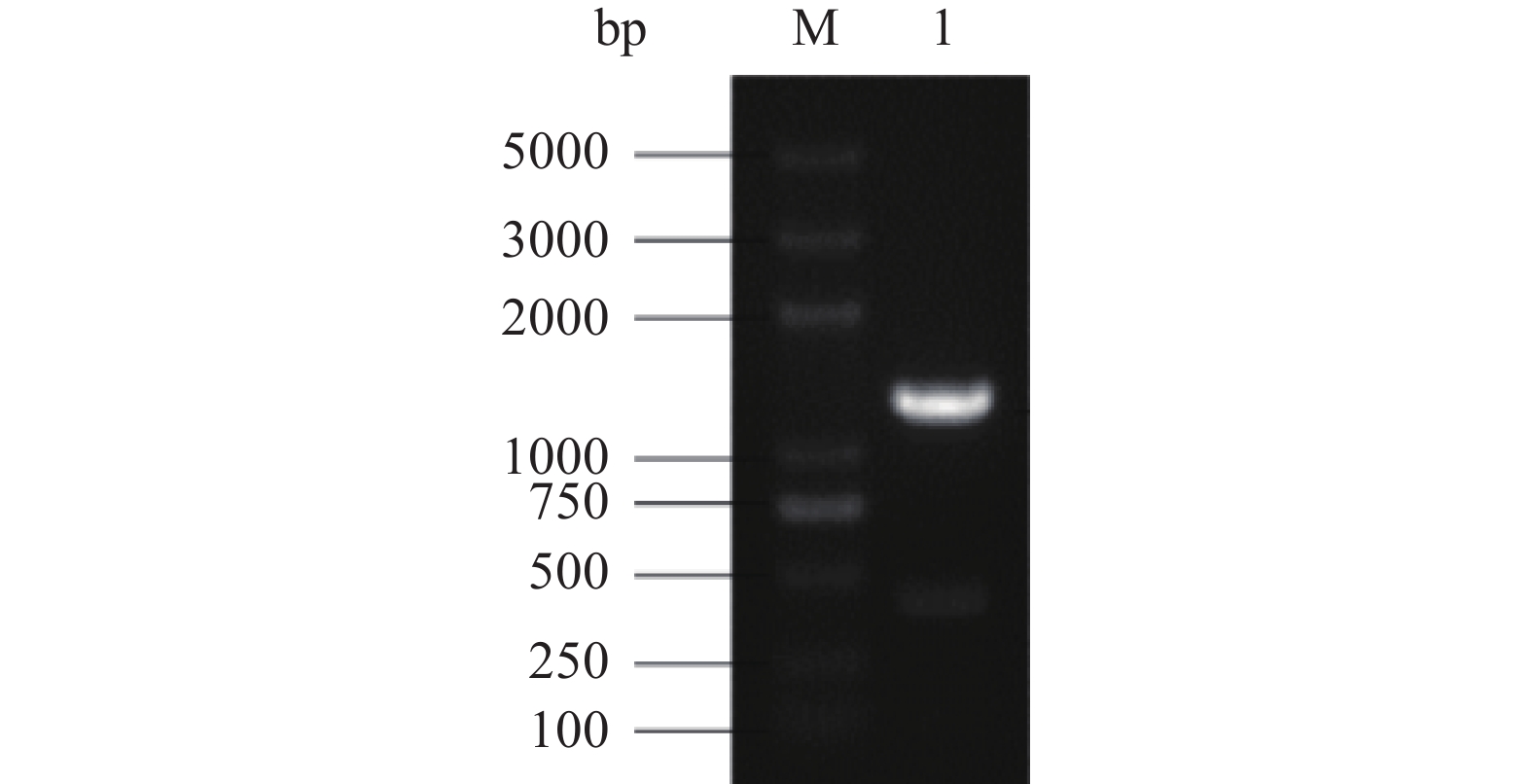
依据黑曲霉pmeA基因组序列(Gene ID: 4980618),应用软件DNAMAN8.0设计用于扩增去除分泌信号肽编码序列的pmeA基因引物P1、P2。提取黑曲霉果胶酶生产菌的基因组,用引物P1、P2进行PCR扩增,基因组提取实验步骤和PCR扩增实验步骤参照文献[21],回收约1300 bp的目的条带,克隆测序,将测序正确的质粒命名为pMD-pmeA。
1.2.2 表达载体的构建
用XbaⅠ和HindⅢ双酶切质粒pMD-PmeA,回收约1300 bp的pmeA基因片段。用NheⅠ和HindⅢ双酶切实验室保存的pSZHG6R载体,回收载体片段。用T4 DNA Ligase将这两个片段23 ℃连接2 h,随后将其转化到大肠杆菌DH5α中,过夜培养,挑取单菌落提质粒,XbaⅠ和HindⅢ双酶切鉴定表达载体pSZHG6R-pmeA是否构建成功。
1.2.3 冻融法转化农杆菌
用冻融法将表达载体pSZHG6R-pmeA转入农杆菌AGL1中[22-23],用引物P3、P2进行菌落PCR鉴定。
1.2.4 黑曲霉转化和纯合转化子鉴定
取匀浆后的新鲜黑曲霉TH-2菌丝与活化后的农杆菌AGL1(pSZHG6R-pmeA)菌液各150 μL混合,涂布在贴玻璃纸的固体PDA平板培养基上,30 ℃恒温培养。待长出菌落,将玻璃纸及其上菌落翻转倒扣在添加300 μg/mL潮霉素的 PDA平板上进行筛选。30 ℃培养1 d后,将玻璃纸揭下,继续30 ℃恒温培养。待上述培养皿长出菌落,挑取至添加300 μg/mL潮霉素的 PDA液体培养基中,30 ℃静止培养3~6 d,取菌丝提取基因组DNA。以TH-2基因组DNA为模版,用引物P3、P4进行PCR扩增,将PCR产物用HindⅢ酶切后进行琼脂糖凝胶电泳[22-23]。获得的纯合重组菌株命名为TH-2(glaA::pmeA)。
1.2.5 asaA基因的敲除
首先根据同源重组的原理敲除TH-2(glaA::pmeA)中的pyrG基因。将黑曲霉TH-2(glaA::pmeA)与农杆菌AGL1(pSZH-∆pyrG)共培养。筛选培养基为含5-FOA和尿苷的CD培养基,用引物P5、P6进行鉴定,其他方法和步骤同1.2.4。获得的纯合重组菌株命名为TH-2(glaA::pmeA∆pyrG)。
然后根据同源重组的原理敲除TH-2(glaA::pmeA∆pyrG)中的asaA基因。将黑曲霉TH-2(glaA::pmeA∆pyrG)与农杆菌AGL1(pSZHA-pyrG)进行共培养。筛选培养基为CD培养基,用引物P7、P8进行鉴定,其他方法和步骤同1.2.4。获得的纯合重组菌株命名为TH-2(glaA::pmeA∆pyrGasaA::pyrG)。
1.2.6 SDS-PAGE
取新鲜的黑曲霉菌株,接种于100 mL液体发酵培养基(玉米浆20 g/L,豆饼粉30 g/L,葡萄糖100 g/L)中,30 ℃,200 r/min培养。将3~9 d的发酵上清液10 μL与10 μL的2×Protein loading buffer混合,沸水浴5~8 min使其中蛋白质变性。以Protein marker作为对比,将3~9 d发酵上清液进行SDS-PAGE检测,浓缩胶电压为100 V,分离胶电压为120 V。电泳结束后用考马斯亮蓝过夜染色,后用脱色液脱色4~6 h。
1.2.7 果胶甲酯酶酶活的检测
取3~9 d重组菌株TH-2(glaA::pmeA)或TH-2(glaA::pmeA∆pyrG∆asaA)的发酵上清液测定果胶甲酯酶酶活。酶活定义为:在50 ℃,pH8.5的条件下,1 mL酶液每分钟催化果胶水解生成甲醛的µmol数为一个酶活力单位,即1 U。具体检测方法参照文献[18]。
1.2.8 果胶甲酯酶酶学性质的检测
将重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)进行发酵培养,取第6 d的发酵上清液,在pH区间3~8.5范围内,以0.5为梯度逐一进行果胶甲酯酶酶活检测,对比不同pH条件下,果胶甲酯酶酶活的变化情况。在温度区间为20~80 ℃范围内,以10 ℃为梯度逐一进行果胶甲酯酶酶活检测,对比不同温度条件下,果胶甲酯酶酶活的变化情况。
2. 结果与分析
2.1 果胶甲酯酶pmeA基因的克隆
提取果胶酶生产菌的基因组,用基因特异性引物P1、P2克隆出1267 bp的果胶甲酯酶基因pmeA如图1所示。将此条带与pMD-19T连接并转入大肠杆菌DH5α中,提取大肠杆菌质粒。将测序得到的质粒序列进行Blast比对,结果表明此序列与Gene ID: 4980618的序列相似性为100%。
2.2 表达载体pSZHG6R-pmeA的构建
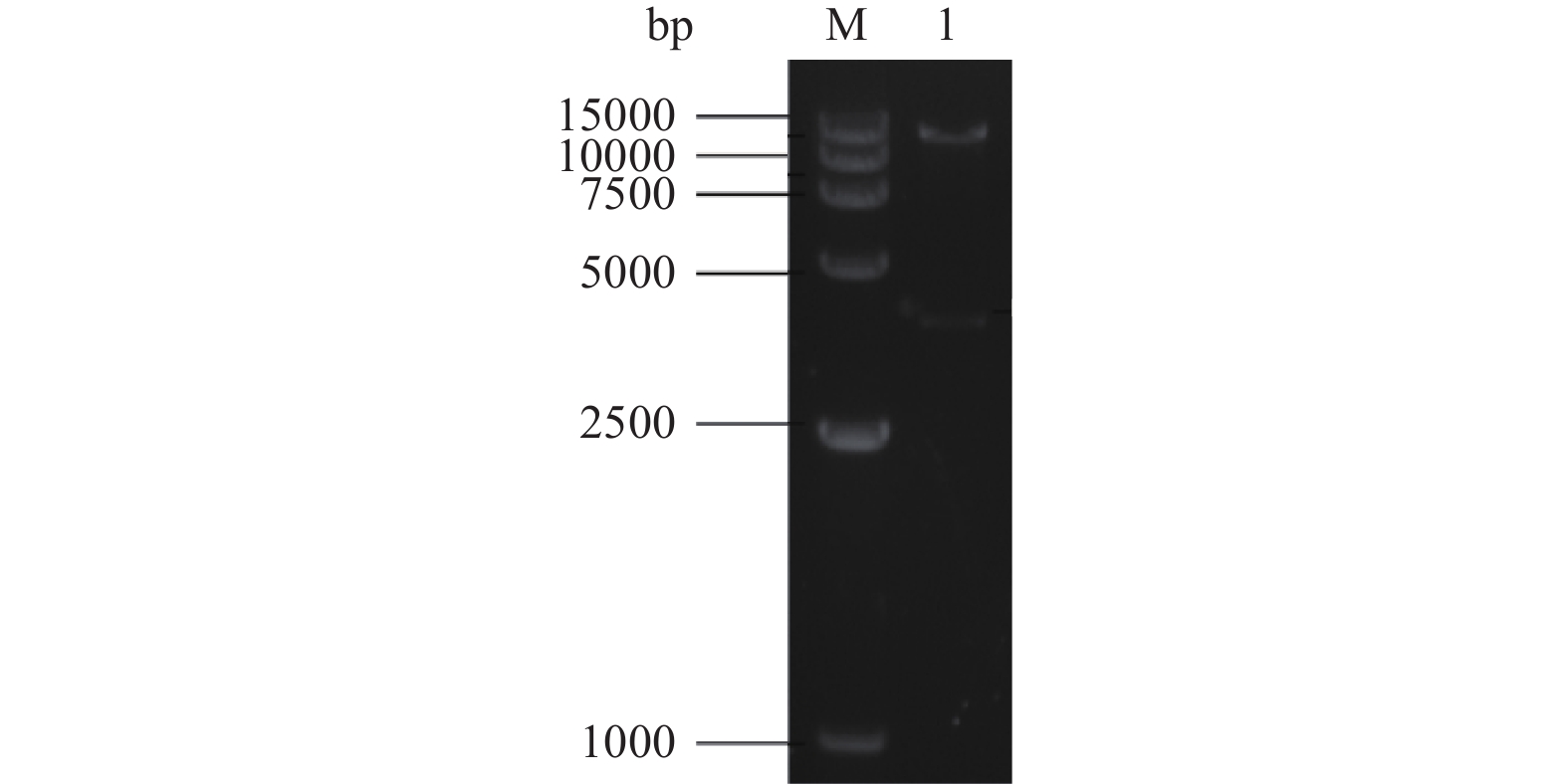
用基因特异性引物P1、P2克隆出1347 bp的果胶甲酯酶基因pmeA。回收目的基因并与pMD-19T载体相连,转入大肠杆菌受体中,进行大肠杆菌质粒的提取,得到质粒pMD-pmeA,并用限制性内切酶XbaⅠ和HindⅢ双酶切鉴定,可切出大小为1347 bp和2700 bp的片段,将双酶切回收的pmeA基因片段与pSZHG6R载体连接。提取转化子的质粒,用XbaⅠ和HindⅢ对其进行双酶切,经琼脂糖凝胶电泳鉴定结果如图2所示,1号泳道显示出2条条带,分别约15000 bp和4000 bp,证明表达载体pSZHG6R-pmeA构建成功。
![]() 图 2 pSZHG6R-pmeA双酶切鉴定结果注:M:DL15000 Maker;1:pSZHG6R-pmeA双酶切鉴定。Figure 2. Identification results of pSZHG6R-pmeA double digestion
图 2 pSZHG6R-pmeA双酶切鉴定结果注:M:DL15000 Maker;1:pSZHG6R-pmeA双酶切鉴定。Figure 2. Identification results of pSZHG6R-pmeA double digestion2.3 冻融法转化农杆菌
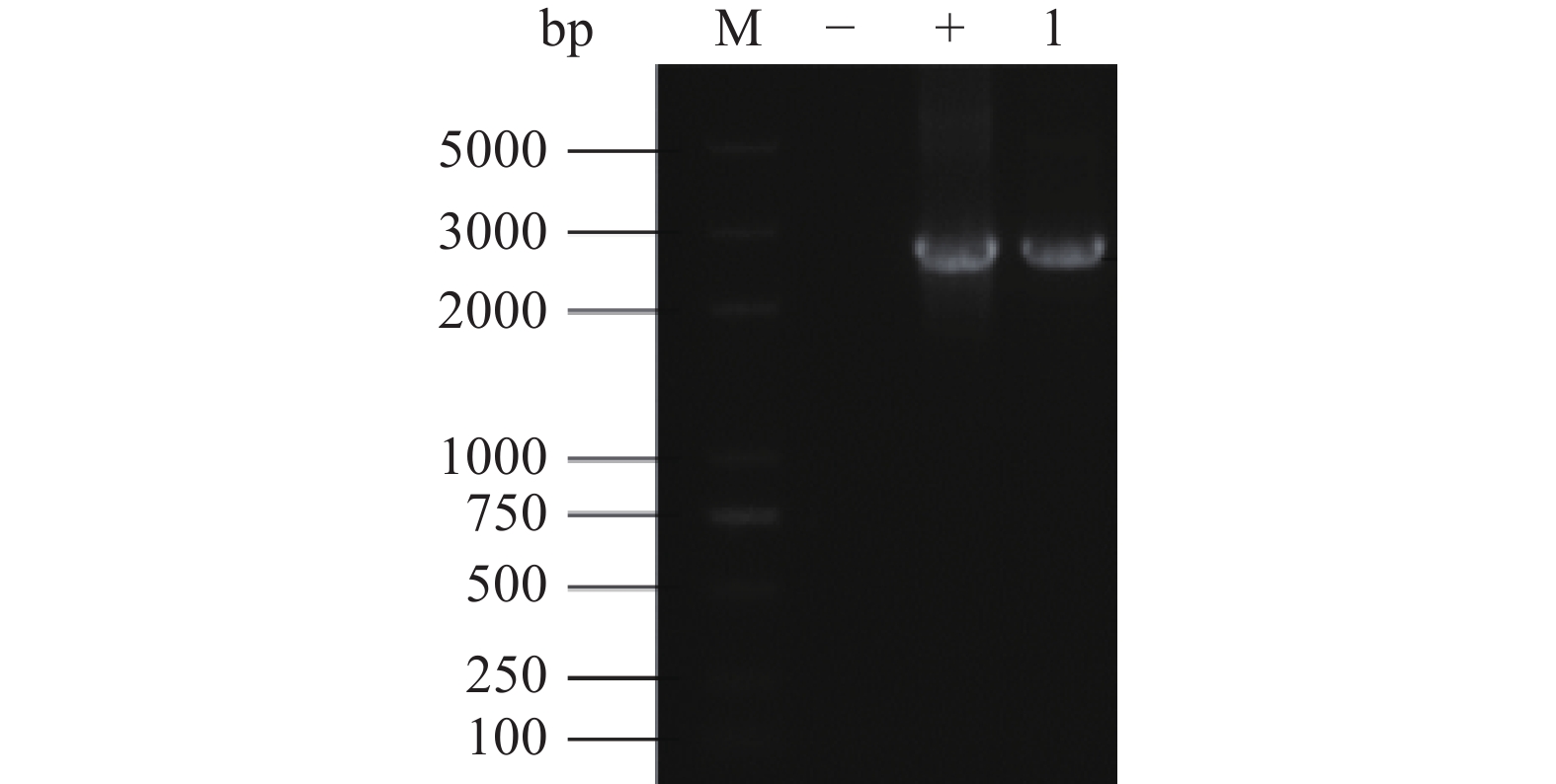
表达载体pSZHG6R-pmeA冻融法转化农杆菌的菌落PCR鉴定结果如图3所示,1号泳道与质粒pSZHG6R-pmeA的阳性对照泳道扩增条带大小相同,证明表达载体pSZHG6R-pmeA成功转入农杆菌中。
![]() 图 3 表达载体pSZHG6R-pmeA转化农杆菌的菌落PCR结果注:M:DL5000 Maker;−:水的阴性对照;+:质粒的阳性对照;1:样本条带。Figure 3. Colony PCR results of expression vector pSZHG6R-pmeA transformed into Agrobacterium tumefaciens
图 3 表达载体pSZHG6R-pmeA转化农杆菌的菌落PCR结果注:M:DL5000 Maker;−:水的阴性对照;+:质粒的阳性对照;1:样本条带。Figure 3. Colony PCR results of expression vector pSZHG6R-pmeA transformed into Agrobacterium tumefaciens2.4 过表达pmeA纯合转化子的筛选和鉴定
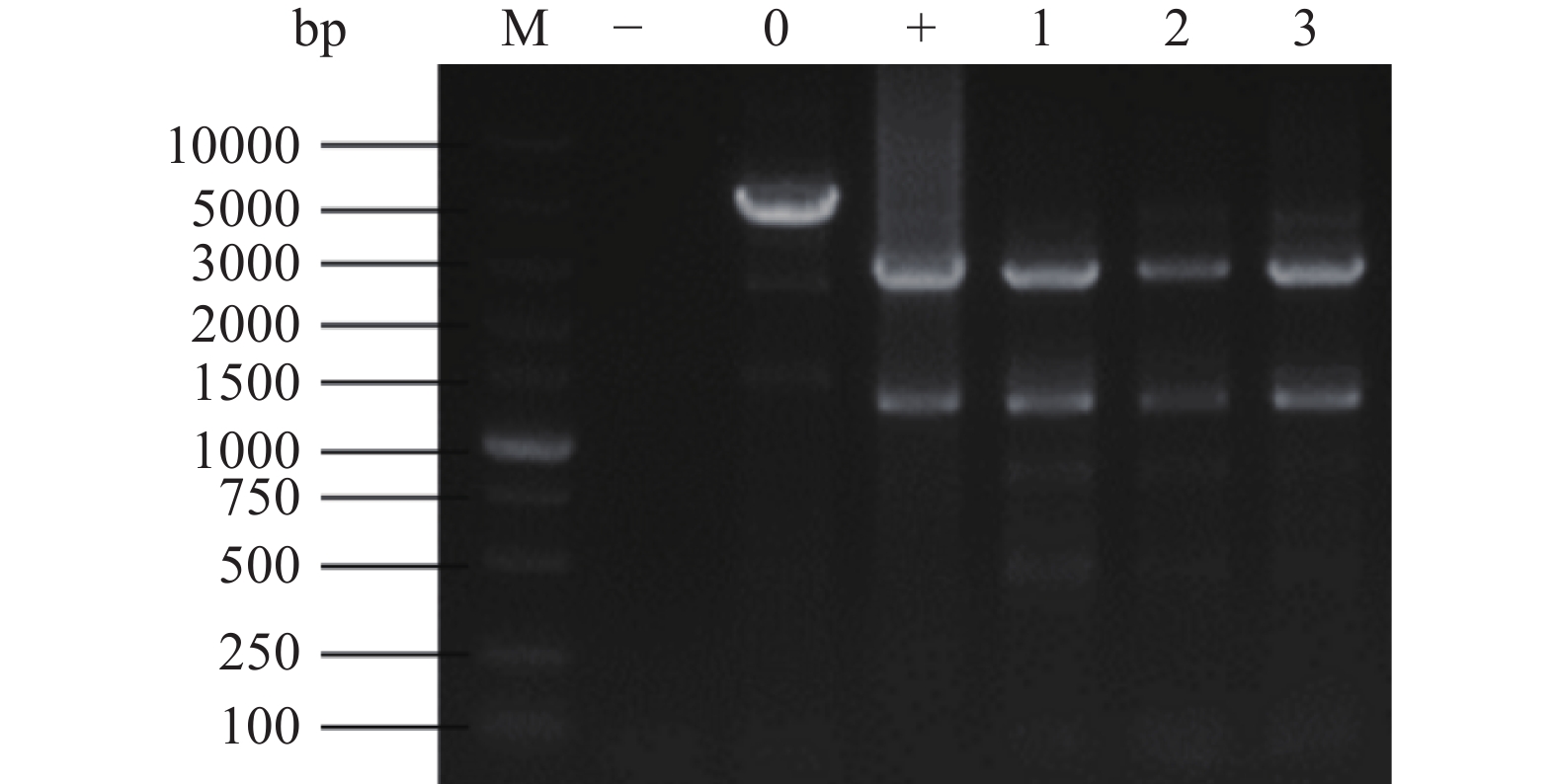
以筛选到的黑曲霉菌株基因组DNA为模版,用引物P3、P4进行PCR扩增,将PCR产物用HindⅢ酶切后进行琼脂糖凝胶电泳,结果如图4所示,1~3号重组菌株可获得与阳性对照质粒pSZHG6R-pmeA相同的条带。所以1~3号重组菌株均为纯合转化子,命名为TH-2(glaA::pmeA)。
![]() 图 4 黑曲霉重组转化子的PCR酶切鉴定结果注:M:DL10000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~3:样本条带。Figure 4. Identification results of PCR digestion of recombinant transformants of Aspergillus niger
图 4 黑曲霉重组转化子的PCR酶切鉴定结果注:M:DL10000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~3:样本条带。Figure 4. Identification results of PCR digestion of recombinant transformants of Aspergillus niger2.5 重组菌株TH-2(glaA::pmeA)的发酵与检测
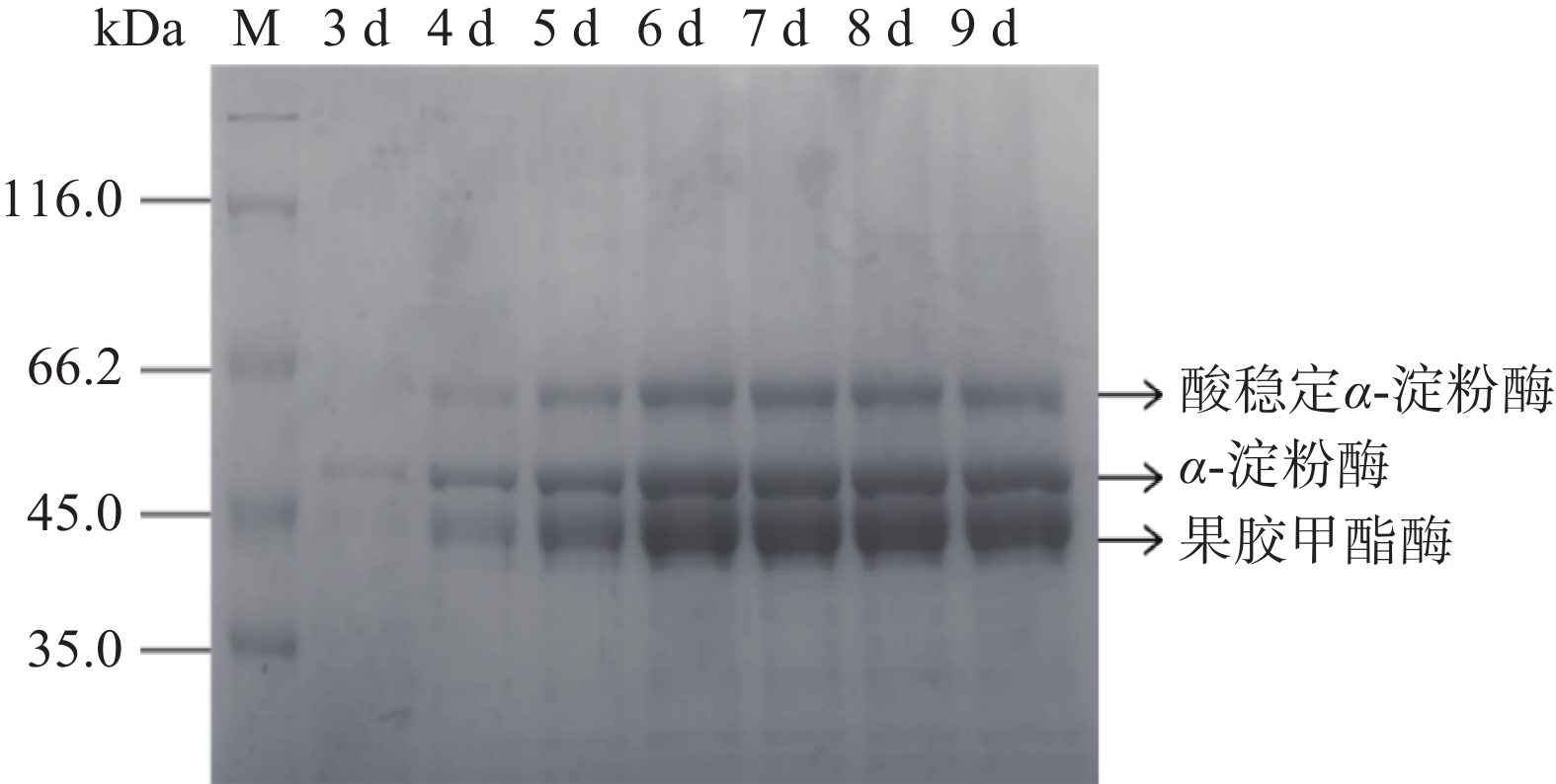
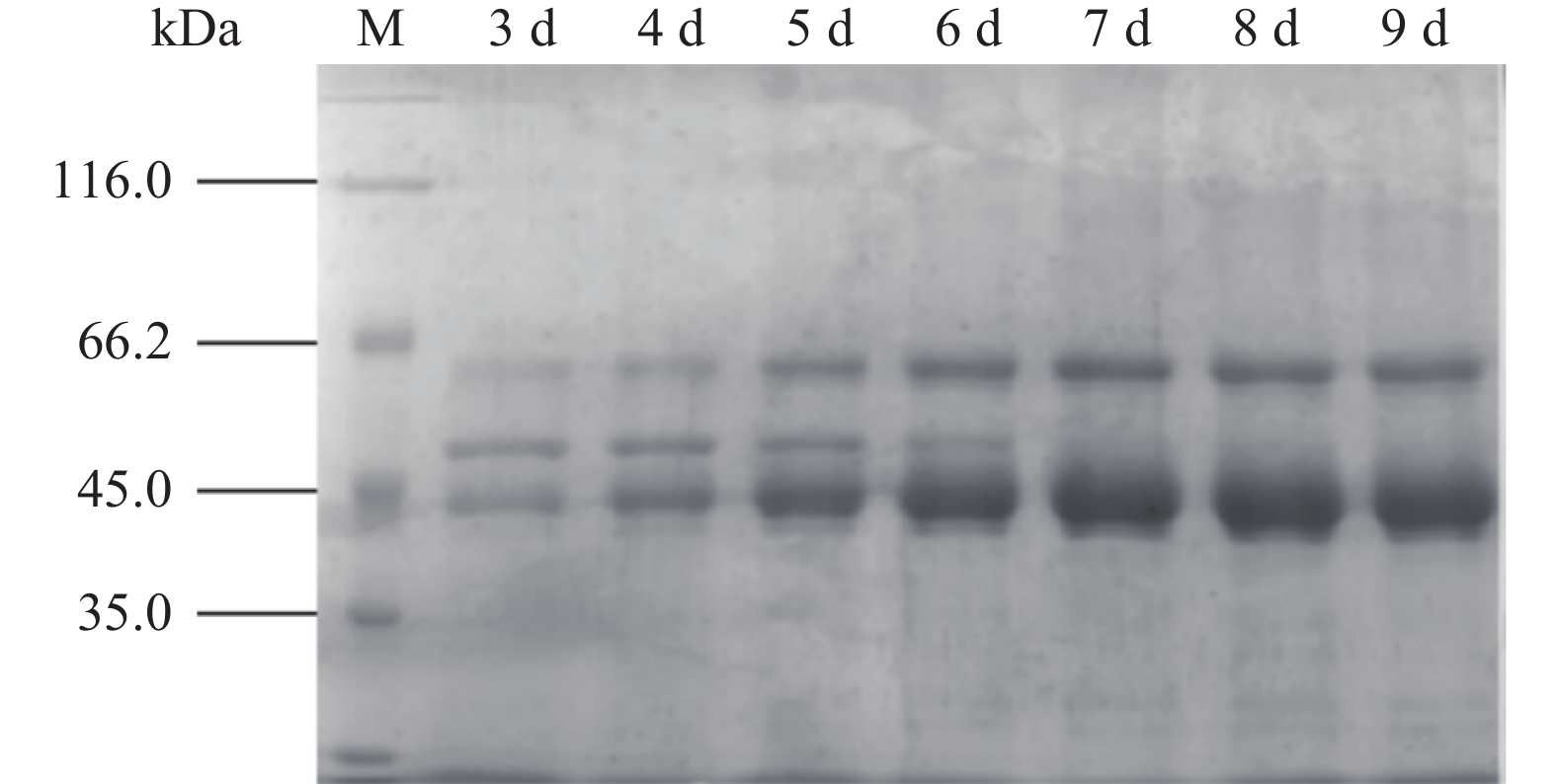
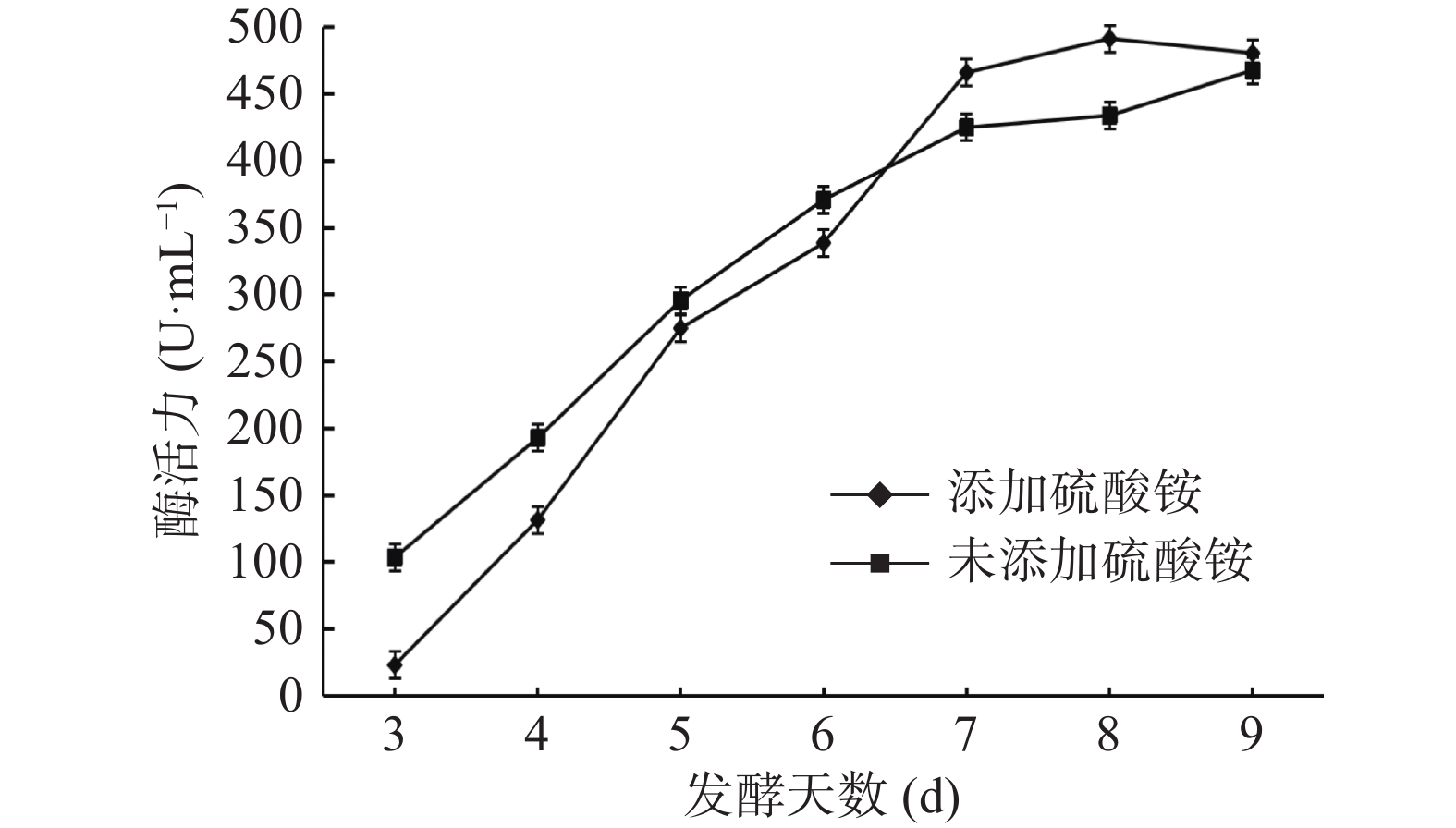
黑曲霉重组菌株TH-2(glaA::pmeA)发酵液上清SDS-PAGE结果如图5所示,在液体发酵培养基中添加1%的硫酸铵后,SDS-PAGE结果如图6所示。由图5可知,52 kDa的α-淀粉酶条带和60 kDa的酸稳定的α-淀粉酶条带为重组菌株的背景蛋白,但在45 kDa处增加了一条明显的蛋白条带,与预测的PmeA分子量34 kDa不符。NetNGlyc 1.0 Server分析结果表明,在PmeA中有4个潜在的N糖基化位点,推断是N糖基化修饰增大了PmeA的分子量。发酵上清液酶活检测结果如图7所示,重组菌株发酵第9 d酶活达到467.77 U/mL。
![]() 图 5 基本发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果Figure 5. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in basic fermentation medium
图 5 基本发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果Figure 5. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in basic fermentation medium![]() 图 6 添加硫酸铵发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果Figure 6. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in fermentation medium with ammonium sulfate
图 6 添加硫酸铵发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果Figure 6. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in fermentation medium with ammonium sulfate![]() 图 7 重组菌株TH-2(glaA::pmeA)的酶活检测结果Figure 7. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA)
图 7 重组菌株TH-2(glaA::pmeA)的酶活检测结果Figure 7. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA)由图5可知,加入硫酸铵的TH-2(glaA::pmeA)发酵样本α-淀粉酶条带逐渐消失。由图7可知,加入硫酸铵后酶活与对照样本相比略有提升,其发酵第8 d酶活最高,达到491.44 U/mL。
2.6 重组菌株TH-2(glaA::pmeA)中pyrG基因的敲除
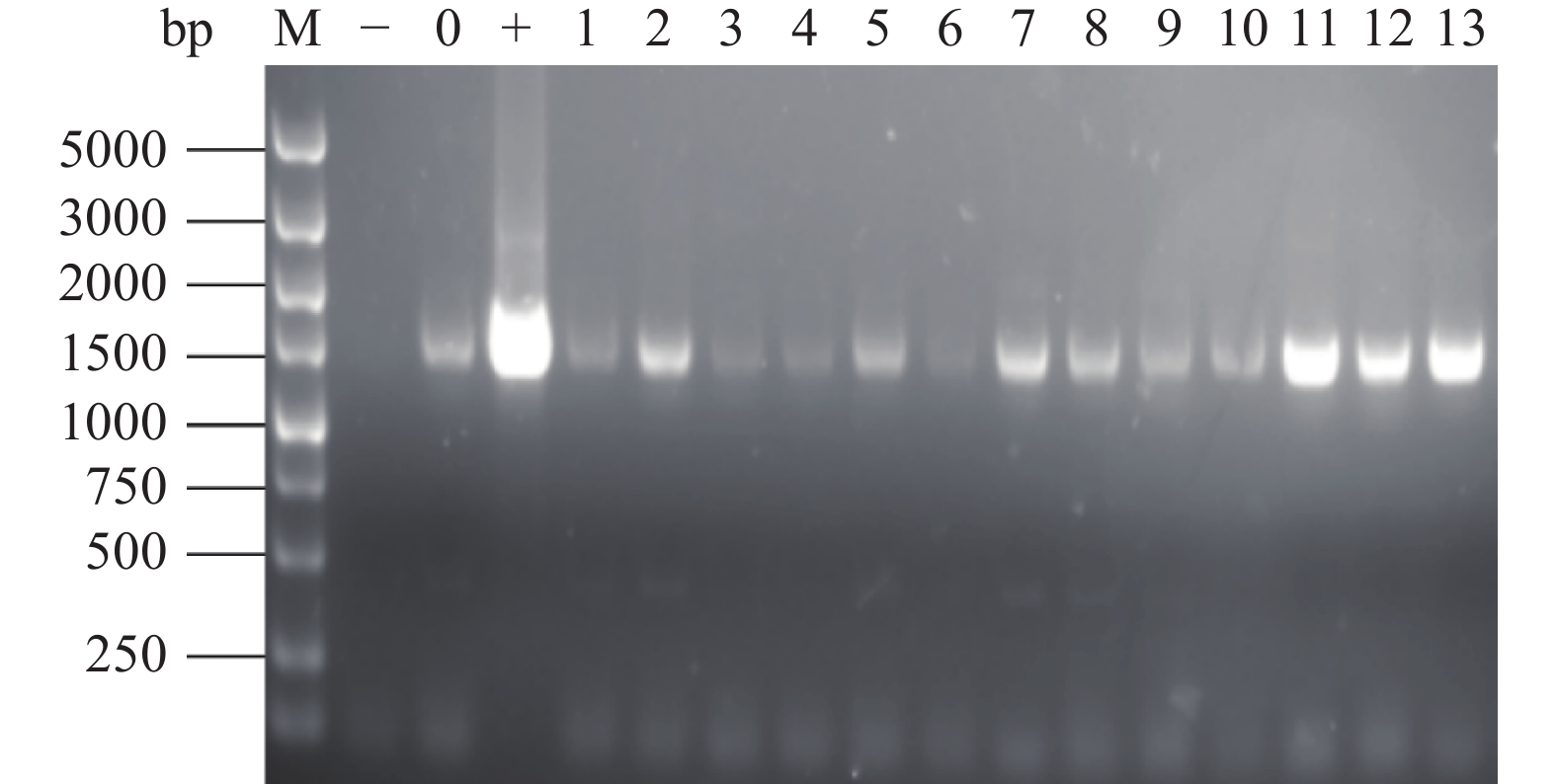
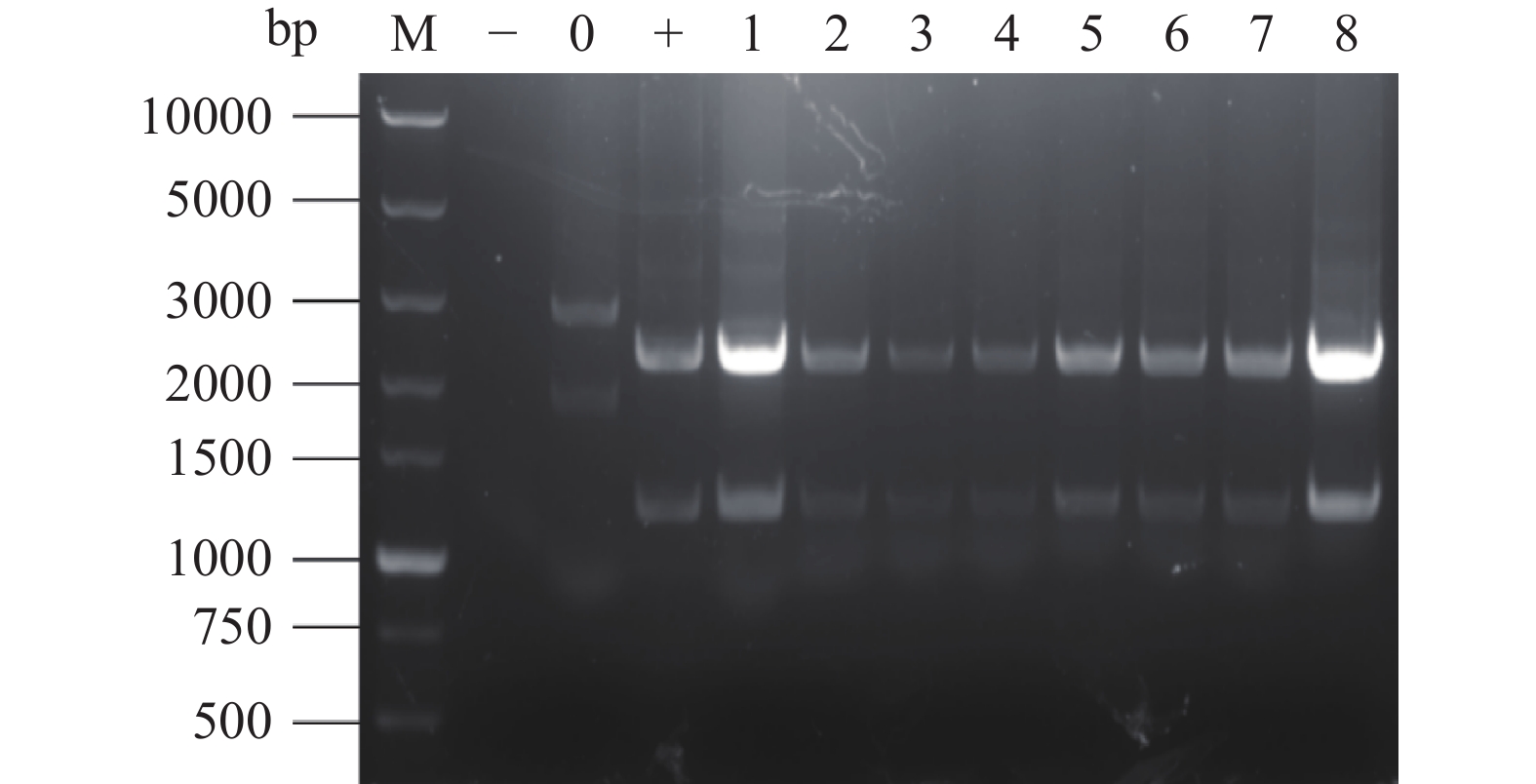
将含敲除原位pyrG基因载体的农杆菌AGL1(pSZH-∆pyrG)与黑曲霉工程菌TH-2(glaA::pmeA)进行共培养,提取转化子的基因组DNA进行PCR扩增和HindIII酶切鉴定,结果如图8所示。1~10号样本出现400 bp左右条带,与阴性对照条带相同,为未转化的出发菌株TH-2(glaA::pmeA)。11、12、13号样本与阳性对照质粒pSZH-∆pyrG的电泳条带相同,均未出现TH-2菌株中的400 bp条带,证明为敲除pyrG基因的纯合转化子,命名为TH-2(glaA::pmeA∆pyrG)。
![]() 图 8 敲除pyrG基因的重组转化子PCR鉴定结果注:M:DL5000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~13:样本条带。Figure 8. PCR identification results of recombinant transformants knocked out of pyrG gene
图 8 敲除pyrG基因的重组转化子PCR鉴定结果注:M:DL5000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~13:样本条带。Figure 8. PCR identification results of recombinant transformants knocked out of pyrG gene2.7 重组菌株TH-2(glaA::pmeA∆pyrG)中asaA基因的敲除
使用本实验室已构建好的敲除asaA基因的农杆菌AGL1(pSZHA-pyrG)与黑曲霉工程菌TH-2(glaA::pmeA∆pyrG)进行共培养。提取转化子的基因组DNA进行PCR扩增和HindIII酶切鉴定。结果如图9所示,1~8号重组菌株与阳性对照质粒pSZHA-pyrG的电泳条带相同,证明为敲除asaA基因的纯合转化子,命名为TH-2(glaA::pmeA∆pyrG∆asaA)。
![]() 图 9 敲除asaA基因的重组转化子PCR酶切鉴定结果(HindIII)注:M:DL10000Maker;-:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~8:样本条带。Figure 9. Identification results of PCR digestion of recombinant transformants knocked out asaA gene(HindIII)
图 9 敲除asaA基因的重组转化子PCR酶切鉴定结果(HindIII)注:M:DL10000Maker;-:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~8:样本条带。Figure 9. Identification results of PCR digestion of recombinant transformants knocked out asaA gene(HindIII)2.8 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)的发酵与检测
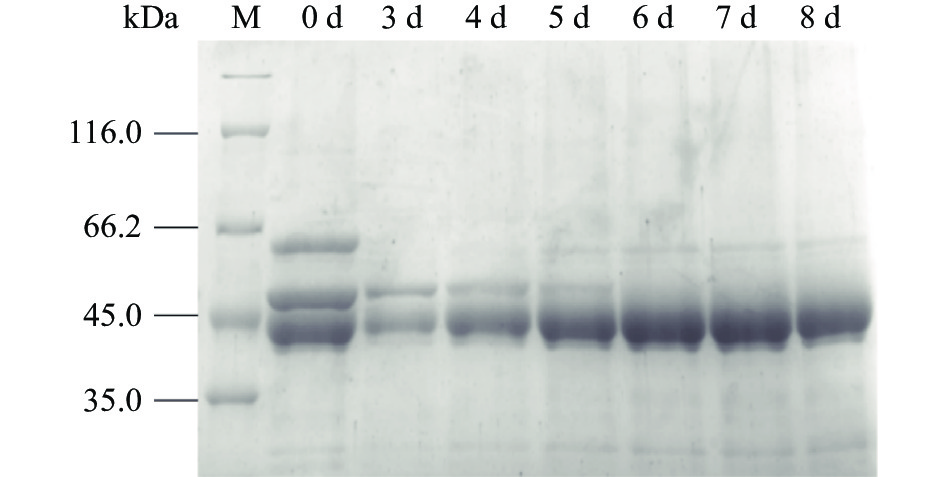
将上述重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)在添加1%硫酸铵的发酵培养基中发酵培养,并进行SDS-PAGE检测,结果如图10所示。在发酵第6 d后可以得到一条明显的果胶甲酯酶蛋白条带,重组菌株TH-2(glaA::pmeA∆pyrG∆asAA)不再分泌60 kDa的酸稳定α-淀粉酶,同时通过加入硫酸铵降解了其分泌的52 kDa的α-淀粉酶背景蛋白。
![]() 图 10 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE检测结果Figure 10. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE test results
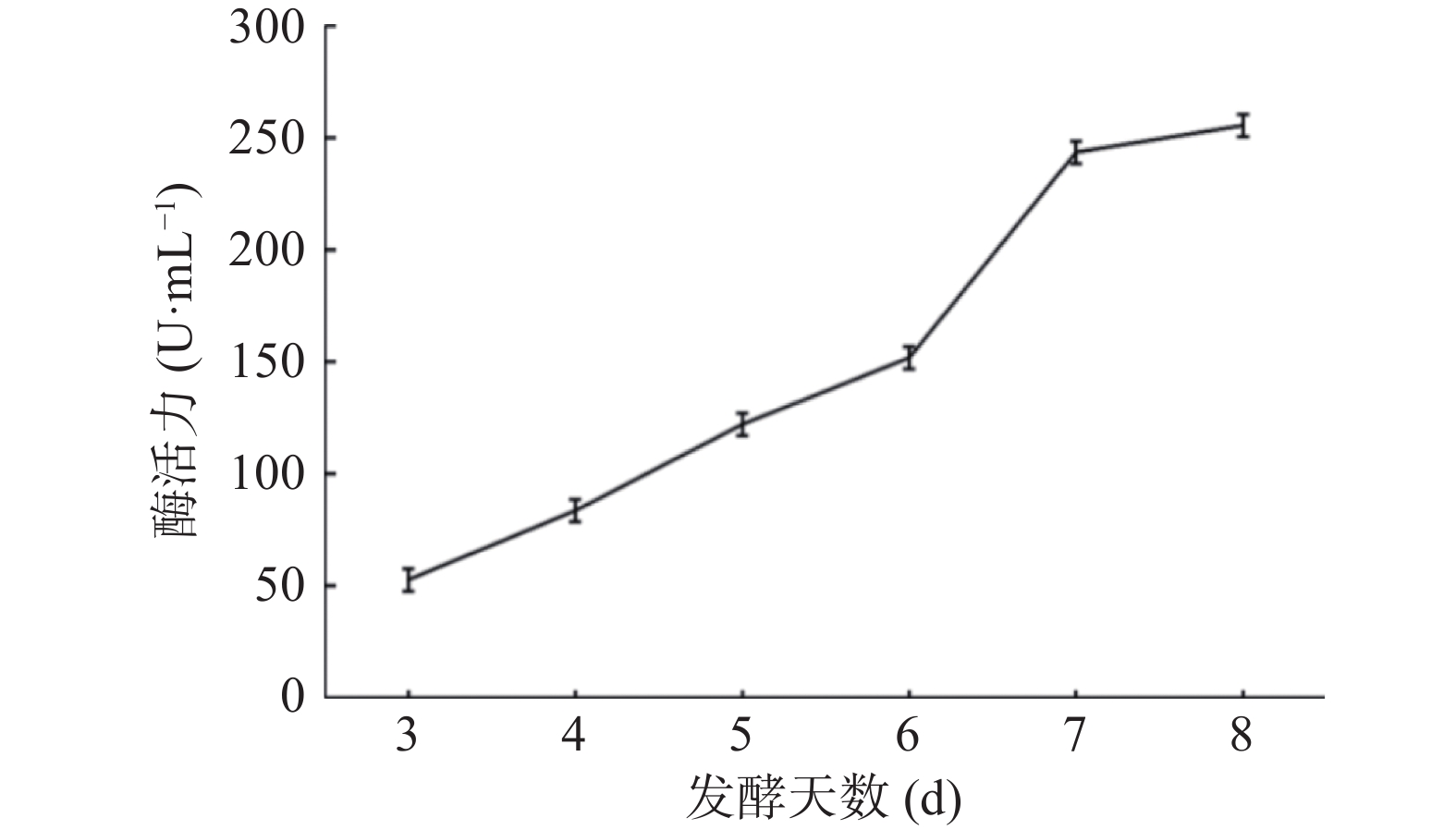
图 10 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE检测结果Figure 10. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE test results取1号纯合重组菌株3~8 d上清发酵液进行酶活测定,结果如图11所示,第8 d酶活最高,可达255.40 U/mL,可以表明1号纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)分泌的果胶甲酯酶具有较高酶活。
![]() 图 11 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)的酶活检测结果Figure 11. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)
图 11 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)的酶活检测结果Figure 11. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)2.9 重组果胶甲酯酶的酶学性质测定
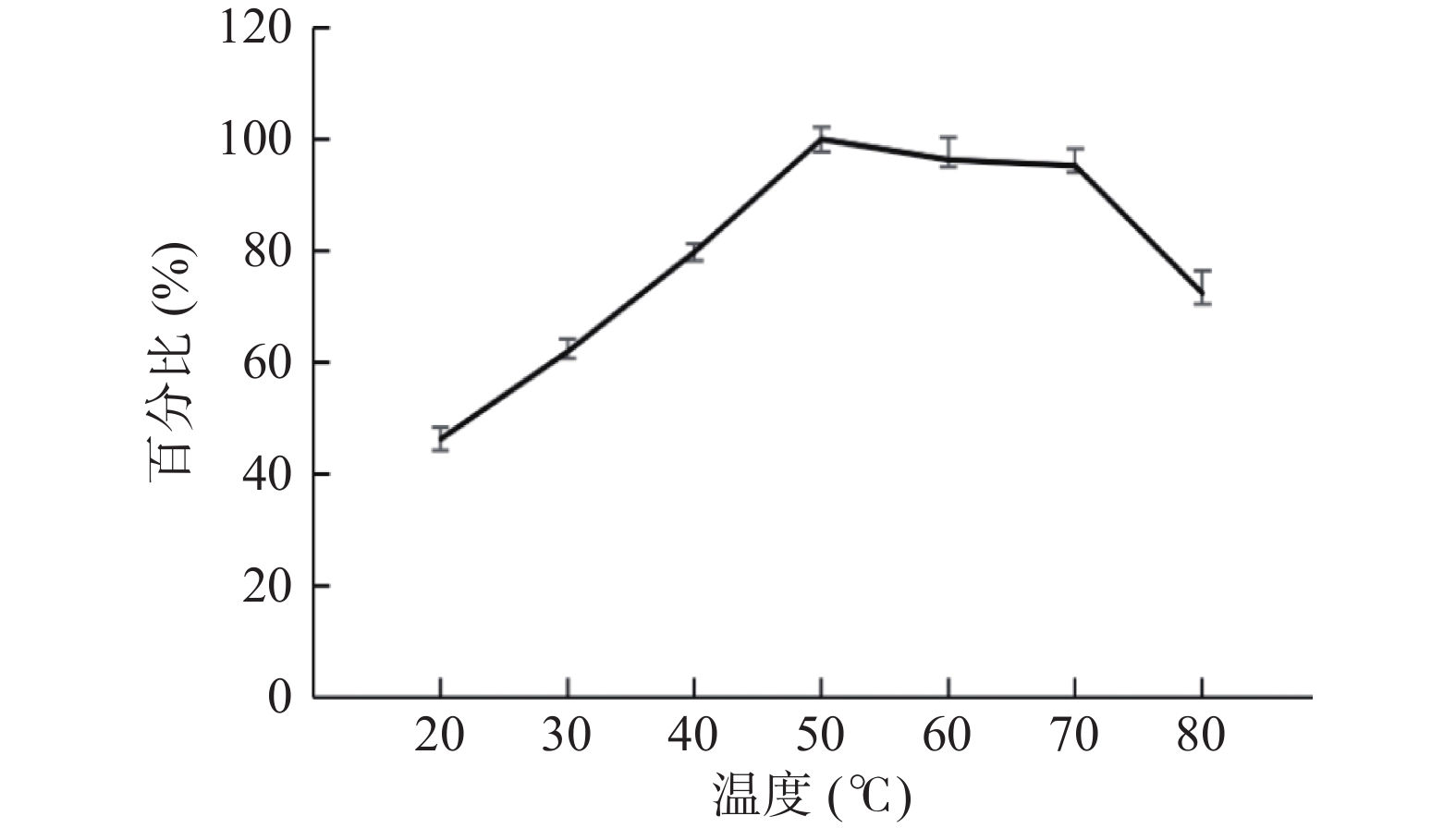
取纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)发酵第8 d上清液。对其在不同温度条件下的酶活进行测定,以50 ℃时的酶活为计算标准,结果如图12所示,在50 ℃时果胶甲酯酶活性到达峰值,随后逐渐回落。其中作用温度为50 ℃时的酶活为263.85 U/mL,作用温度为80 ℃时的酶活仍能达到191.24 U/mL,比在常温20 ℃条件下的酶活性还要高。可以得出结论,纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)分泌的果胶甲酯酶的最适作用温度为50 ℃,并具有较好的耐热性,在80 ℃高温条件下仍能维持其酶活性的70%以上。
![]() 图 12 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同温度的酶活检测结果Figure 12. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different temperatures
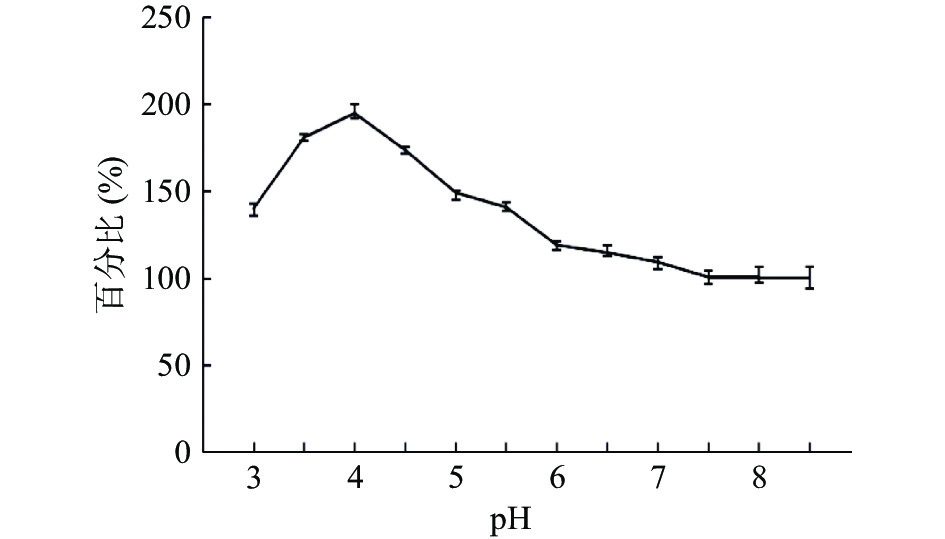
图 12 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同温度的酶活检测结果Figure 12. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different temperatures取纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)分泌的第6 d发酵上清液。对其在不同pH条件下的酶活进行测定,以pH为8.5时酶活为计算标准,结果如图13所示,其中作用pH为4时的酶活最高,最高酶活为397.50 U/mL,为pH为8.5时酶活的195%。随后酶活随pH增加呈逐渐下降趋势。可以得出结论,纯合重组菌株TH-2(glaA::PmeA∆pyrG∆asaA)分泌的果胶甲酯酶的最佳pH作用范围是3.0~5.0之间,最适作用pH为4.0。
![]() 图 13 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同pH的酶活检测结果Figure 13. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different pH
图 13 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同pH的酶活检测结果Figure 13. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different pH3. 讨论与结论
本实验成功构建果胶甲酯酶的超量表达载体,并将其转入受体菌TH-2中,获得纯合重组菌株TH-2(glaA::pmeA)。在发酵培养基中加入硫酸铵,可以有效消除背景蛋白中的α-淀粉酶。可能因为硫酸铵被菌体利用后,使得发酵培养基pH下降[24-26],而菌体在发酵后期也会分泌酸性物质,最终导致发酵后期培养基pH下降幅度更大,激活了胞外的酸性蛋白酶活性,使α-淀粉酶被分解[27-28],赵波等[29]为控制酵母高密度发酵过程中pH的变化,在发酵液中添加硫酸铵取得成效。但是在此条件下,背景蛋白中的酸稳定α-淀粉酶未被分解。进一步敲除了酸稳定α-淀粉酶基因获得纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA),通过发酵时加入硫酸铵,获得纯度较高的果胶甲酯酶[30-31]。但测得其最高发酵酶活为255.40 U/mL,低于同样条件下的黑曲霉工程菌TH-2(glaA::pmeA)的酶活491.44 U/mL,可能是asAA基因或pyrG基因的敲除对菌体的生长和代谢产生了不利影响,其原因还有待于进一步研究。
重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)分泌的果胶甲酯酶在50 ℃条件下酶活性最高,最高酶活达263.85 U/mL;并且在80 ℃的高温条件下,仍能保持酶活性的70%以上。其最适作用的pH范围是3.0~5.0,最佳作用pH为4.0,在pH4.0时,最高酶活达397.50 U/mL。所以,纯合重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)分泌的果胶甲酯酶在酸性条件下酶促效率更高;此外,在pH为7.0时,其酶活性为最适作用pH时的60%左右。表明用本研究构建的工程菌生产的果胶甲酯酶具有较宽的作用温度和pH范围,具有很好的应用前景。
-
![]()
图 2 pSZHG6R-pmeA双酶切鉴定结果
注:M:DL15000 Maker;1:pSZHG6R-pmeA双酶切鉴定。
Figure 2. Identification results of pSZHG6R-pmeA double digestion
![]()
图 3 表达载体pSZHG6R-pmeA转化农杆菌的菌落PCR结果
注:M:DL5000 Maker;−:水的阴性对照;+:质粒的阳性对照;1:样本条带。
Figure 3. Colony PCR results of expression vector pSZHG6R-pmeA transformed into Agrobacterium tumefaciens
![]()
图 4 黑曲霉重组转化子的PCR酶切鉴定结果
注:M:DL10000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~3:样本条带。
Figure 4. Identification results of PCR digestion of recombinant transformants of Aspergillus niger
![]()
图 5 基本发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果
Figure 5. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in basic fermentation medium
![]()
图 6 添加硫酸铵发酵培养基重组菌株TH-2(glaA::pmeA)SDS-PAGE检测结果
Figure 6. SDS-PAGE detection results of recombinant strain TH-2(glaA::pmeA)in fermentation medium with ammonium sulfate
![]()
图 7 重组菌株TH-2(glaA::pmeA)的酶活检测结果
Figure 7. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA)
![]()
图 8 敲除pyrG基因的重组转化子PCR鉴定结果
注:M:DL5000 Maker;−:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~13:样本条带。
Figure 8. PCR identification results of recombinant transformants knocked out of pyrG gene
![]()
图 9 敲除asaA基因的重组转化子PCR酶切鉴定结果(HindIII)
注:M:DL10000Maker;-:水的阴性对照;0:出发菌株的阴性对照;+:质粒的阳性对照;1~8:样本条带。
Figure 9. Identification results of PCR digestion of recombinant transformants knocked out asaA gene(HindIII)
![]()
图 10 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE检测结果
Figure 10. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)SDS-PAGE test results
![]()
图 11 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)的酶活检测结果
Figure 11. Enzyme activity test results of recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)
![]()
图 12 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同温度的酶活检测结果
Figure 12. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different temperatures
![]()
图 13 重组菌株TH-2(glaA::pmeA∆pyrG∆asaA)不同pH的酶活检测结果
Figure 13. Recombinant strain TH-2(glaA::pmeA∆pyrG∆asaA)enzyme activity test results at different pH
表 1 PCR引物序列
Table 1 PCR Primer sequence
引物名称 引物序列 P1:pmeA-sense TCTAGACGCATGACGGCTCCTT P2:pmeA-antisense AAGCTTAGATCTTAGTTGATGTAGCTAGTATC P3:5’GLA-sense TCTAGACTCGGCGACTTGGTCTTCAC P4:3’GLA-antisense CTCGAGCATAAGGCGGGTTCACATC P5:5’pyrG-sense GAATTCCTCGAGCTAACATACATTCCG P6:3’pyrG-antisense TCTAGAGGTAAGTAATCAGTAACTG P7:5’asaA-sence TCGCCTATCTACTCCTCATCC P8:3’asaA-antisence CTTCAACCCTGCTGCTAAATC  下载: 导出CSV
下载: 导出CSV
-
[1] 王胜, 孟昆, 罗会颖, 等. 果胶甲酯酶的结构与功能研究进展[J]. 生物工程学报,2020,36(6):1021−1030. [Wang Sheng, Meng Kun, Luo Huiying, et al. Research progress on structure and function of pectin methylesterase[J]. Journal of Bioengineering,2020,36(6):1021−1030. [2] 何英东, 崔文甲, 李晓, 等. 果胶甲酯酶的作用机理及在食品加工中的应用[J]. 食品研究与开发,2020,41(12):185−189. [He Ying Dong, Cui Wen Jia, Li Xiao, et al. Mechanism of pectin methylesterase and its application in food processing[J]. Food Research and Development,2020,41(12):185−189. doi: 10.12161/j.issn.1005-6521.2020.12.031 [3] Tao T, Kun M, Huiying L, et al. New insights into the role of T3 Loop in determining catalytic efficiency of GH28 endo-polygalacturonases[J]. Plos One,2015,10(9):e0135413. doi: 10.1371/journal.pone.0135413
[4] 刘化禹, 娄爽, 秦栋, 等. 蓝果忍冬果柄离区形成中内源激素含量与细胞壁相关酶活性的变化特征[J]. 西北植物学报,2019,39(1):115−125. [Liu Huayu, Lou Shuang, Qin Dong, et al. Variation characteristics of endogenous hormone content and cell wall related enzyme activity in the formation of stem abscission zone of Lonicera macranthoides[J]. Acta Botanica Sinica,2019,39(1):115−125. [5] Khubber S, Chaturvedi K, Thakur N, et al. Low-methoxyl pectin stabilizes low-fat set yoghurt and improves their physicochemical properties, rheology, microstructure and sensory liking[J]. Food Hydrocolloids,2020,111:106240.
[6] Ricardo Durán Baron, Churio C J G, Juana Bermúdez. Addition of high esterification pectin as a stabilizer in the production of low fat yogurt[J]. ITEGAM-Journal of Engineering and Technology for Industrial Applications (ITEGAM-JETIA),2020,6(26):13−17.
[7] 贾玉姣, 裴雅琨, 朱玉涛, 等. 植物果胶甲酯酶应答病虫抗性与耐铝胁迫的研究进展[J]. 分子植物育种, 2021,19(12):4127-4132. Jia Yujiao, Pei Yakun, Zhu Yutao, et al. Research progress of plant pectin methylesterase in response to disease and insect resistance and aluminum stress[J]. Molecular Plant Breeding, 2021,19(12):4127-4132.
[8] Kent Lisa M, Loo Trevor S, Melton Laurence D, et al. Structure and properties of a non-processive, salt-requiring, andacidophilic pectin methylesterase from Aspergillus niger provide insights into the key determinants of processivity control[J]. The Journal of Biological Chemistry,2015:M115.673152.
[9] Fontana R C, Silveira M M. Influence of pectin, glucose, and pH on the production of endo- and exo-polygalacturonase by Aspergillus oryzae in liquid medium[J]. Brazilian Journal of Chemical Engineering,2012,29(4):683−690. doi: 10.1590/S0104-66322012000400001
[10] Jiang X, Chen P, Yin M, et al. Constitutive expression, purifification and characterisation of pectin methylesterase from Aspergillus niger in Pichia pastoris for potential application in the fruit juice industry[J]. Journal of the Science of Food and Agriculture,2013,93:375−381. doi: 10.1002/jsfa.5771
[11] 李晓, 王文亮, 王月明, 等. 外源性果胶甲酯酶对低盐腌渍黄瓜质构性质的影响[J]. 食品科学技术学报,2018,36(6):88−94. [Li Xiao, Wang Wenliang, Wang Yueming, et al. Effect of exogenous pectin methylesterase on texture properties of low salt pickled cucumber[J]. Journal of Food Science and Technology,2018,36(6):88−94. doi: 10.3969/j.issn.2095-6002.2018.06.013 [12] Gordon E Anthon, Diane M Barrett. Pectin methylesterase activity and other factors affecting pH and titratable acidity in processing tomatoes[J]. Food Chemistry,2012,132(2):915−920. doi: 10.1016/j.foodchem.2011.11.066
[13] 王志云, 罗会颖, 姚斌, 等. Bispora sp. MEY-1来源嗜酸果胶甲酯酶性质分析及其应用研究[J]. 中国农业科技导报,2020(5):60−70. [Wang Zhiyun, Luo Huiying, Yao bin, et al. Characterization and application of eosinophilic pectin methylesterase from bispora sp. mey-1[J]. China Agricultural Science and Technology Guide,2020(5):60−70. [14] 阎然, 杨晓颖, 傅茂润, 等. 外源果胶甲酯酶和氯化钙复合处理对树莓采后品质的影响[J]. 食品与发酵工业,2019,45(18):130−136. [Yan Ran, Yang Xiaoying, Fu Maorun, et al. Effect of exogenous pectin methylesterase and calcium chloride on postharvest quality of raspberry[J]. Food and Fermentation Industry,2019,45(18):130−136. [15] 毛小龙. 黑曲霉ANSTJ01菌株安全性评价及鸡球虫EtMIC2外源蛋白表达[D]. 泰安: 山东农业大学, 2018. Mao Xiaolong. Safety evaluation of Aspergillus niger ANSTJ01 and expression of Eimeria ETMIC2 foreign protein[D]. Taian: Shandong Agricultural University, 2018.
[16] Rajulapati V, Dhillon A, Gali K K, et al. Green bioprocess of degumming of jute fibers and bioscouring of cotton fabric by recombinant pectin methylesterase and pectate lyases from Clostridium thermocellum[J]. Process Biochemistry,2020,92:93−104. doi: 10.1016/j.procbio.2020.02.024
[17] Li Y, He H, He L F. Genome-wide analysis of the pectin methylesterase gene family in potato[J]. Potato Research,2020:1−19.
[18] 焦云鹏. 商品果胶酶中果胶酯酶的分离纯化[J]. 中国酿造,2008(20):74−76. [Jiao Yunpeng. Separation and purification of pectinesterase in commercial pectinase[J]. China Brewing,2008(20):74−76. [19] 李菀, 胡月, 李光霞, 等. 纤维素酶和果胶酶提取对甘草渣多糖抗氧化和抗肿瘤性能的影响[J]. 食品工业科技,2020,41(9):309−313319. [Li Wan, Hu Yue, liguangxia, et al. The effect of cellulase and pectinase extraction on the antioxidation and antitumor properties of Glycyrrhiza residue polysaccharide[J]. Food Industry Science and Technology,2020,41(9):309−313319. [20] Jérô me Pelloux, Christine Rustérucci, Mellerowicz E J. New insights into pectin methylesterase(PME) structure and function[J]. Trends in Plant Science,2007,12(6):267−277. doi: 10.1016/j.tplants.2007.04.001
[21] 张会. β-1, 4-内切木聚糖酶基因在黑曲霉中安全高效表达的研究[D]. 哈尔滨: 东北农业大学, 2013. Zhang Hui. Study on the safe and efficient expression of β-1, 4-endoxylanase gene in Aspergillus niger[D]. Harbin: Northeast Agricultural University, 2013.
[22] 李杰, 张贺, 王欣, 等. 多拷贝脂肪酶基因在黑曲霉中表达研究[J]. 东北农业大学学报,2018,49(4):49−58. [Li Jie, Zhang He, Wang Xin, et al. Expression of multi copy lipase gene in Aspergillus niger[J]. Journal of Northeast Agricultural University,2018,49(4):49−58. doi: 10.3969/j.issn.1005-9369.2018.04.006 [23] 李杰, 张会, 张莹莹, 等. 食品级木聚糖酶黑曲霉工程菌的构建[J]. 东北农业大学学报,2013,44(11):7−13. [Li Jie, Zhang Hui, Zhang Yingying, et al. Construction of food grade xylanase engineered Aspergillus niger[J]. Journal of Northeast Agricultural University,2013,44(11):7−13. doi: 10.3969/j.issn.1005-9369.2013.11.002 [24] 刘军, 张云鹏, 冯治平, 等. 赖氨酸发酵废渣中硫酸铵的去除[J]. 粮食与饲料工业,2009(6):36−37. [Liu Jun, Zhang Yunpeng, Feng Zhiping, et al. Removal of ammonium sulfate from lysine fermentation residue[J]. Food and Feed Industry,2009(6):36−37. doi: 10.3969/j.issn.1003-6202.2009.06.012 [25] 娄百勇, 宋祺, 曹华伟, 等. 不同无机盐浓度对金霉素发酵生产影响[J]. 科技创新与应用,2018(27):43−46. [Lou Baiyong, Song Qi, Cao Huawei, et al. Effects of different inorganic salt concentrations on chlortetracycline fermentation[J]. Innovation and Application of Science and Technology,2018(27):43−46. [26] Sipriyadi S, Wahyudi A T, Suhartono M T, et al. Optimization of xylanase production by Streptomyces costaricanus 45I-3 using various substrates through submerged fermentation[J]. Microbiology Indonesia,2020,14(1):5. doi: 10.5454/mi.14.1.5
[27] AnthonG. E, Barrett D. M. Comparison of threecolorimetric reagents for the determination of methanol with alcohol oxidase: Application to the assay of pectin methylesterase[J]. Journal of Agricultural and Food Chemistry,2004,52:3749−3753. doi: 10.1021/jf035284w
[28] 王娟, 张汝玲, 王红红, 等. 头孢菌素C发酵过程中硫酸铵加量对菌丝形态及效价的影响[J]. 求医问药(下半月),2011,9(7):40. [Wang Juan, Zhang ruling, Wang Honghong, et al. Effect of ammonium sulfate dosage on mycelial morphology and potency during cephalosporin C fermentation[J]. Seeking Medical Advice (second half),2011,9(7):40. [29] 赵波, 赵文杰, 顾敏, 等. 硫酸铵与pH对酵母高密度发酵生产谷胱甘肽的影响[J]. 中国医药工业杂志,2009(3):180−182. [Zhao Bo, Zhao Wenjie, Gu min, et al. Effects of ammonium sulfate and pH on glutathione production by yeast high density fermentation[J]. Chinese Journal of Pharmaceutical Industry,2009(3):180−182. doi: 10.3969/j.issn.1001-8255.2009.03.008 [30] 王晴, 胡智慧, 钱玉梅, 等. 柑橘果胶酯酶分离条件及生物学特性研究[J]. 廊坊师范学院学报(自然科学版),2020,20(1):69−71, 85. [Wang Qing, Hu Zhizhi, Qian Yumei, et al. Study on separation conditions and biological characteristics of pectinesterase from Citrus[J]. Journal of Langfang Normal University (Natural Science Edition),2020,20(1):69−71, 85. [31] 王华芳, 陈玮. 酶法提取苹果渣果胶工艺研究[J]. 三门峡职业技术学院学报,2020,19(1):135−139. [Wang Huafang, Chen Wei. Study on enzymatic extraction of pectin from apple pomace[J]. Journal of Sanmenxia Vocational and Technical College,2020,19(1):135−139. doi: 10.3969/j.issn.1671-9123.2020.01.023
 下载:
下载:













计量
- 文章访问数: 351
- HTML全文浏览量: 159
- PDF下载量: 30
