


Research Progress on the Mechanism of Lactic Acid Bacteria in Improving Diabetes Metabolism
-
摘要: 乳酸菌对糖尿病人有良好的治疗功效,其机制的探究也越发深入。乳酸菌可抑制肠道中α-葡萄糖苷酶使血糖降低,经由调节胰岛素信号传导、糖代谢、脂质代谢等相关代谢途径来降低血糖水平,通过促进胰岛素的表达或降低磷酸化水平改善胰岛素抵抗,影响机体内的葡萄糖转运因子进而从糖的重吸收等方面影响葡萄糖的代谢,下调脂肪合成酶的表达以抑制脂肪生成,同时改善肠道菌群的丰度并影响肠道菌群的完整性对炎症反应和代谢紊乱进行修复。本文对目前研究乳酸菌改善糖尿病的具体作用机制进行研究论述,为未来乳酸菌预防和治疗糖尿病提供参考。Abstract: Lactic acid bacteria have a good therapeutic effect on diabetic patients, and the exploration of its mechanism has been intensified. Lactic acid bacteria can inhibit α-glucosidase in the intestine to reduce blood sugar, and reduce blood sugar levels by regulating insulin signal transduction, sugar metabolism, lipid metabolism and other related metabolic pathways. Promote insulin expression or reduce phosphorylation level to improve insulin resistance and affect the body. The glucose transport factors in turn affects glucose metabolism from the aspects of sugar reabsorption, down-regulates the expression of liposynthetase, inhibits adipogenesis, improves the abundance of intestinal flora and affects the integrity of intestinal flora, and affects inflammation and metabolic disorders to repair it. This article reviews the current research on the specific mechanism of lactic acid bacteria in improving diabetes, and provides references for the future prevention and treatment of diabetes by lactic acid bacteria.
-
Keywords:
- lactic acid bacteria /
- diabetes mellitus /
- hypoglycemia /
- mechanism
-
糖尿病是一类胰岛素分泌缺陷或障碍引起的代谢性疾病,分为I型和II型。目前世界患有糖尿病的人数逐年增加,据国际糖尿病联盟(IDF)数据,2017年全球糖尿病患者约有4.25亿人,其中约80%的患者集中在中低收入国家,预计到2045年患病人数可能达到6.29亿[1]。在这些病患中,中国占1.14亿并且有逐年上涨的趋势[2-3],这些患者中II型糖尿病(T2DM)患者占90%以上。因此,糖尿病的预防和治疗具有重要的意义。T2DM主要症状为胰岛素分泌不足、胰岛素敏感性降低或者两者皆有[4],其次为脂肪、蛋白质、水、电解质代谢紊乱。现在应用较为广泛的降糖药物主要为双胍类、促胰岛素分泌类(包括磺酰脲类和非磺酰脲类)、噻唑烷二酮类、葡萄糖苷酶抑制剂等。然而传统治疗药物仍然具有很大的局限性,例如肿胀、胀气、腹泻和继发性衰竭、治疗效果不理想等[5],因此继续寻找或开发新的降糖药物是目前急需解决的一个重要问题。
乳酸菌是一类能分解碳水化合物产生大量乳酸的细菌统称。乳酸菌分布广泛,种类繁多,按照其形态分为球菌和杆菌,按生长温度可分为高温型菌和中温型菌。按来源可分为动物源与植物源乳酸菌,按照发酵类型可分为异型发酵菌和同型发酵菌[6]。作为一种益生菌,乳酸菌功能丰富,具有增强机体免疫功能、降低胆固醇、缓解乳糖不耐受、预防癌症、维持肠道微生态平衡等功能[7-9]。研究发现,通过摄入乳酸菌能够抑制α-葡萄糖苷酶[10]、丰富肠道菌群[11]、调节代谢的途径[12]从而发挥治疗糖尿病的作用。乳酸菌作为一种天然产物,具有副作用小的优势,降血糖乳酸菌的开发为未来治疗和预防糖尿病能够提供一种新的思路,对未来减少糖尿病患者的数量有着积极的作用。
本文主要从乳酸菌抑制α-葡萄糖苷酶活性[10]、调节胰岛素信号传导通路、糖代谢通路、脂代谢通路[12]、改善肠道菌群的丰度和完整性[11]等方面进行论述。通过探讨乳酸菌对II型糖尿病造成的代谢损伤进行改善修复,以期为未来研究乳酸菌改善糖尿病造成的机体损伤和预防糖尿病提供新的参考和思路。
1. 对α-葡萄糖苷酶的抑制作用
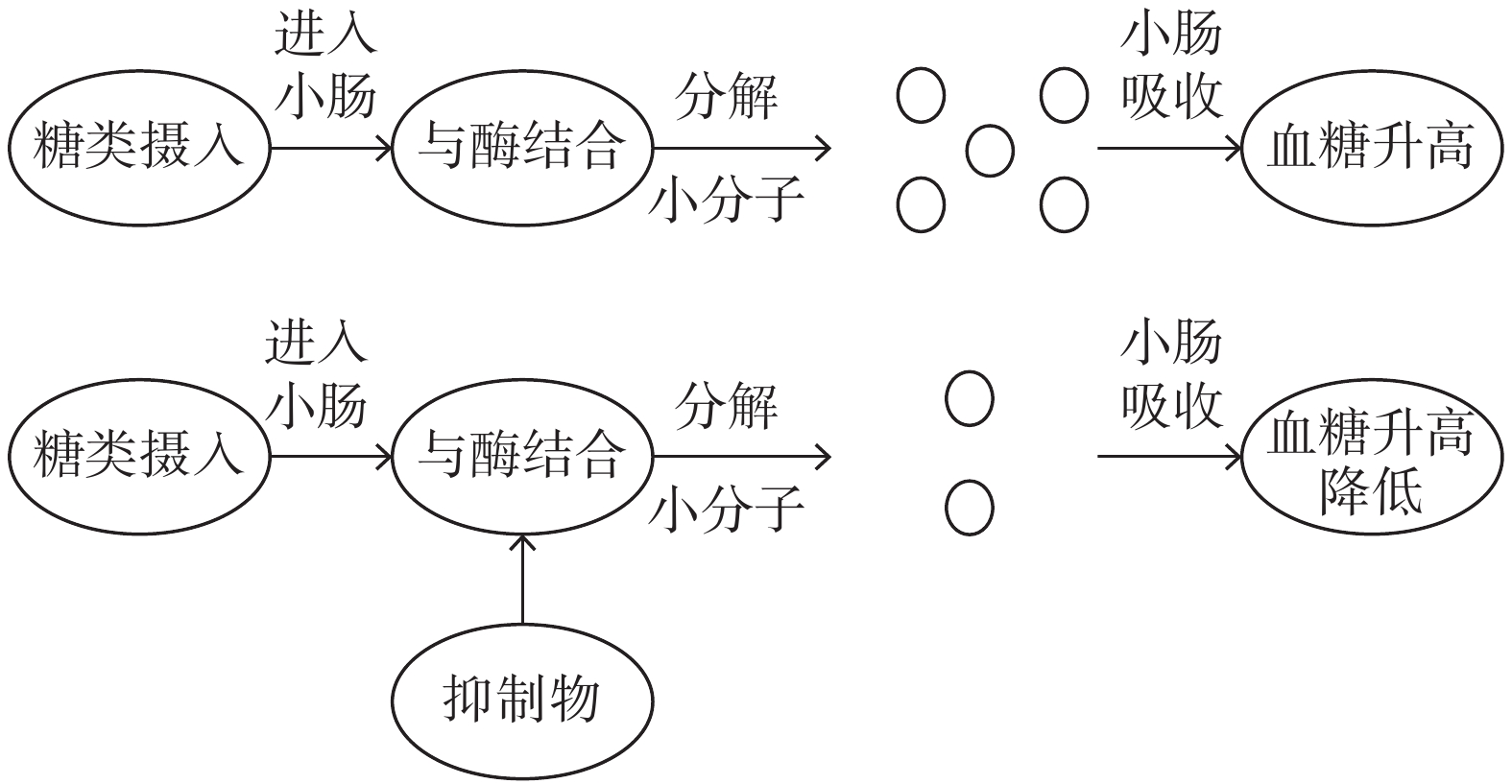
在糖尿病的治疗中,以抑制α-葡萄糖苷酶为主要的治疗方法,其具体机制为α-葡萄糖苷酶抑制剂能够竞争性地与α-葡萄糖苷酶上的位点相结合,使摄入的食物中糖类分解降低,使多余的糖类物质直接被排入大肠,从而降低了餐后血糖,见图1。目前治疗糖尿病的药物中主要作用机制为抑制α-葡萄糖苷酶。例如米格列醇(Miglitol)、阿卡波糖(Acarbose)和伏格列波糖(Voglibose)等[13],但是药物的副作用很多,而乳酸菌引起的副作用少,具有明显的优势。
![]() 图 1 抑制α-葡萄糖苷酶降低血糖原理Figure 1. Principle of inhibiting α-glucosidase to lower blood sugar
图 1 抑制α-葡萄糖苷酶降低血糖原理Figure 1. Principle of inhibiting α-glucosidase to lower blood sugarα-葡萄糖苷酶是一类能分解碳水化合物的关键酶统称,广泛分布在小肠各段上,主要位于小肠粘膜细胞刷状缘上,在糖代谢中占据重要地位,筛选降血糖乳酸菌时其对α-葡萄糖苷酶的抑制作用是重要参考指标。通过抑制α-葡萄糖苷酶能控制糖类进入体内代谢循环,降低血糖值。它的抑制与糖代谢相关。但目前研究中筛选降血糖乳酸菌是通过体外对α-葡萄糖苷酶抑制能力来进行判别是否具有降血糖功能。因此研究α-葡萄糖苷酶的抑制能力十分重要。Li等[14]从车前草中分离出植物乳杆菌对α-葡萄糖苷酶抑制能力达到32.19%,并且在随后的研究结果中证明通过体外对乳酸菌α-葡萄糖苷酶抑制能力进行筛选后,其在治疗糖尿病中也表现出良好的效果。副干酪乳杆菌对糖尿病模型小鼠有良好的治疗效果,并且能够对生化指标的进行调节,对α-葡萄糖苷酶的抑制能力达到31.89%[15]。因此,抑制α-葡萄糖苷酶的能力是降血糖乳酸菌筛选的重要指标,也是乳酸菌治疗糖尿病的作用机制之一。
2. 乳酸菌对代谢通路的改善
糖尿病是一种代谢疾病,会导致多器官的并发症。其对各种代谢途径均会产生影响,主要为胰岛素信号转导通路、糖代谢通路、脂质代谢通路。而乳酸菌的摄入能通过影响其信号通路进而对糖尿病造成的损伤起到改善作用。
2.1 乳酸菌摄入对胰岛素信号传导通路的调节
胰岛素信号通路的功能障碍可能导致代偿性高胰岛素血症,从而产生胰岛素抵抗。乳酸菌能够对胰岛素信号传导通路中的相关分子进行调控,对相关通路的信号分子进行激活,促进胰岛素分泌,并对胰岛素抵抗进行改善,恢复胰岛素的代谢功能[16]。近些年研究了乳酸菌对胰岛素信号传导的调节作用,对乳酸菌治疗影响胰岛素抵抗的具体机制进行了探讨。
Li等[17]研究发现口服酪乳杆菌可以通过PI3K/Akt信号通路调节胰岛素抵抗,通过介导PI3K/Akt信号通路的激活,从而减轻T2DM大鼠的胰岛素抵抗。进一步研究摄入乳酸菌对胰岛素信号通路的调节,Korkmaz等[18]发现摄入乳酸菌有效地促进了IRS-1、eNOS和AKT的蛋白表达,然后对胰岛素代谢通路进行了调节,证明了乳酸菌的摄入能调节胰岛素信号传导途径。Sumlu等[19]研究发现饮食中的高果糖通过降低大鼠的IRS-1/2、PI3K、AKT和eNOS的表达来抑制肝胰岛素信号传导,而植物乳杆菌的补充可以逆转肝脏IRS-1/AKT/eNOS信号的减弱,恢复胰岛素信号的传导,瑞士乳杆菌也可以调节IRS-1表达。
胰岛素在细胞内的作用是通过其受体(IR)的激活来启动的,信号通过IRS-1和IRS-2传递给其主要效应器AKT和eNOS[19]。在对模型小鼠的组织器官研究中,IR-1、IRS-2和AKT的表达或磷酸化降低,能够揭示器官中胰岛素抵抗的水平[20]。
2.2 乳酸菌摄入对糖代谢通路的调节
摄入乳酸菌能缓解糖尿病引起的高血糖,具体降低血糖的方式可分为两种。乳酸菌能够通过两种途径对单糖进行分解代谢,即6-磷酸葡萄糖的形式进入糖酵解或莱洛伊尔(Leloir)途径进行代谢;通过影响机体内的葡萄糖转运因子进而从糖的重吸收等方面去影响葡萄糖的代谢。
在最近的研究中发现,肾脏中的特异性葡萄糖转运体SGLT2和特殊果糖转运体GLUT5这两种转运蛋白的表达上调,能使肾小管葡萄糖重吸收增加,进而表明胰岛素抵抗状态下肾胰岛素信号转导和SGLT2、GLUT5表达之间的关系[21]。植物乳杆菌和瑞士乳杆菌的摄入降低了SGLT2表达,限制了葡萄糖的重吸收,进而对血糖进行调节。此外,GSK-3B是一种丝氨酸激酶,在肝糖原合成过程中起重要作用,它能抑制肝糖原合成酶(GS)活性,降低肝糖原合成。因此,抑制gsk-3B可通过增加糖原合成和胰岛素敏感性来改善T2D[22]。已有研究表明,副干酪链球菌能下调GSK-3的表达,增加肝糖原含量,对糖尿病进行治疗[23]。Yadav等[24]研究发现在链脲佐菌素诱导的糖尿病大鼠中,葡萄糖的过度生成可能与肝脏糖异生作用的过度或组织对葡萄糖的利用降低有关。两种糖异生关键酶,即葡萄糖-6-磷酸酶(G6Pase)和磷酸烯醇丙酮酸羧激酶(pepck)的RNA表达在糖尿病患者的肝脏中增加[25]。用益生菌发酵乳治疗后,肝脏中这些酶和葡萄糖转运体的表达明显正常化,因此降低了血液中的葡萄糖水平。
2.3 乳酸菌摄入对脂质代谢通路的调节
糖尿病除了导致高血糖症状外,还能够诱发高脂血症。因此,脂质代谢的调节在糖尿病中起到重要影响。乳酸菌摄入能够通过调节信号通路降低血液中脂质水平,还可以通过调节肠道菌群对脂质代谢通路进行调控。
此前,有研究发现乳酸菌可以降低血液中的总胆固醇和低密度脂蛋白水平,并且对肝脏中的脂肪含量有着改善作用[26]。脂肪细胞分化的主要信号通路为:Wnt信号通路、PPAR通路、SREBP通路、细胞外信号调节激酶/丝裂原活化蛋白激酶(ERK–MAPK)信号通路、单磷酸腺苷/蛋白激酶A(cAMP/PKA)信号通路和Hedgehog信号通路[27]。通过之前的研究发现高糖饮食能够显著诱导大鼠肝脏产生脂质的基因,具体表现为SREBP-1c、FAS和Elvol6的表达上调,而通过摄入具有治疗糖尿病效果的乳酸菌后,基因的表达得到明显的改善,对肝脏中的脂质积聚有抑制作用[28]。SREBP-1c是SREBP家族成员之一,参与脂肪酸代谢过程中酶的合成,如FAS[29]。蛋白水解酶S1P和S2P有助于SREBP前体成熟,调节脂质合成相关基因的表达。而在最近的研究中,发现短链脂肪酸(Short-chain fatty acid,简称SCFAs)对肠道的脂代谢通路有影响,SCFAs中的丁酸能够激活脂代谢相关信号通路[12]。乳酸菌通过调节肠道菌群进而对肝脏中的脂代谢水平进行了调节。路晓杰等[30]研究发现肠道菌含量升高,显著抑制了脂质代谢异常,而通过调整肠道微生物的比例,可显著减少脂肪的堆积,并可普遍缓解脂质代谢异常的病理症状。肠道内微生物可以直接或间接影响脂质代谢,还可以发酵膳食纤维产生SCFAs,在能量和脂质代谢中起着重要的作用[31]。Qian等[32]研究发现乳酸菌具有促进小肠内SCFAs生成,影响肥胖小鼠脂质代谢过程,调节小鼠脂质代谢的作用。Zhao等[33]发现植物乳杆菌的补充增加了因高脂饮食减弱的AMPK磷酸化,随后增加了ACC磷酸化水平。这些数据表明,植物乳杆菌NA136的补充是通过AMPK途径磷酸化ACC,抑制SREBP-1/FAS信号来达到降脂的目的。Park等[34]研究发现益生菌可以改变肠道菌群,而肠道微生物群会显著影响宿主的脂质代谢。益生菌处理导致下调SREBP1、FAS和SCD1的表达,进而对脂肪生成进行抑制(见表1)。
表 1 乳酸菌对代谢通路的调节Table 1. Regulation of metabolic pathways by lactic acid bacteria3. 乳酸菌摄入对肠道菌群的调节
3.1 肠道菌群对炎症修复作用
越来越多的研究证明II型糖尿病与肠道菌群之间存在着联系,而在研究乳酸菌治疗糖尿病对肠道菌群所产生的具体影响时发现,糖尿病和高脂饮食能够使肠道内的SCFAs含量降低,乳酸菌摄入能够通过调节肠道菌群的丰度和含量进而调节SCFAs,恢复肠道内SCFAs水平[35]。
Horie等[35]对比分析了正常小鼠与T2DM小鼠的肠道菌群,发现正常小鼠与糖尿病小鼠的肠道菌群的数量有着明显的差异,这说明糖尿病患者的肠道稳态发生了改变。肠道菌群组成的调节可能有助于SCFAs的转化,进而对炎症反应有良好的治疗效果。研究表明,肠道内的SCFA包括乙酸、丙酸和丁酸[36]。而通过补充乳酸菌,肠道内的产酸菌丰度提高,进而恢复肠道内短链脂肪酸含量,SCFAs对肠道内的炎症因子等有调节作用,因此发挥治疗糖尿病的功能[37]。Wang等[38]通过给糖尿病模型小鼠连续喂食干酪乳杆菌,对模型小鼠的肠道菌群的丰度和数量有着显著的调节作用,并且对其涉及途径研究发现,益生菌可以通过调节益生菌-肠道菌群-丁酸盐-炎症途径来治疗糖尿病造成的危害。通过对罗伊氏乳杆菌[39]的研究也证明了降血糖乳酸菌可以通过调节肠道菌群影响肠道SCFAs的含量,进而对糖尿病造成的损伤起到治疗效果。因此,降血糖乳酸菌可通过调节肠道菌群的丰度、维持肠道微生物的稳态和调节肠道激素的水平,进而对糖尿病造成的损伤进行调节。
3.2 肠道菌群修复调节稳态水平
摄入乳酸菌可以改善肠道菌群的完整性,降低全身内毒素(LPS)水平、增加胰高血糖素样肽-1(GLP-1),降低内质网应激,提高外周胰岛素敏感性,进而对糖尿病造成的损伤起到治疗作用。
研究乳酸菌治疗糖尿病的过程中发现,长期的高脂饮食喂养会导致肠道环境中菌群的降低和LPS水平的升高[40]。在正常情况下,LPS的渗漏是由肠道完整性控制的,而肠道完整性是由紧密连接蛋白(TJPs)维持的。TJPs可防止LPS进入全身循环,但高脂饮食的摄入降低了TJP的表达[41]。而乳酸菌可以通过对肠道完整性的修复从而改善这一情况。另外,研究表明肠道微生物群和多器官功能障碍之间存在着动态的串扰,而某些菌株特异性益生菌可以对代谢紊乱起到有益的作用。通过口服摄入乳酸菌修复肠道菌群,可以治疗糖尿病造成的损伤。这一途径主要是调节了肠道菌群的数量以及种类,恢复正常肠道菌群水平,对体内异常的代谢水平进行调节。
3.3 通过肠道菌群预防糖尿病
高脂饮食的增加能使肠道菌群构成发生变化[42],而肠道内的部分菌能够分泌有害物质对胰岛细胞产生危害。乳酸菌的摄入能够补充肠道菌群的丰度,抑制肠道内有害细菌的生长,对糖尿病起到预防的作用。Dumas等[43]通过研究发现肠道菌群中含有谷氨酸脱羧酶,这说明某些菌具有合成或分泌谷氨酸的能力,而谷氨酸的分泌能够使肠源淋巴细胞破坏胰岛细胞,从而引起糖尿病。而乳酸菌的摄入使肠道菌群得到补充,并且抑制其他菌的生长,从而起到预防糖尿病的作用。
4. 总结
糖尿病是一种代谢疾病,可以影响多器官多途径的代谢。而目前通过乳酸菌改善糖尿病造成的损伤具有良好的效果,随着对乳酸菌改善糖尿病损伤的进一步研究,其具体机制也逐渐被发现。首先乳酸菌能够通过抑制α-葡萄糖苷酶使进入体内的小分子糖含量降低,从而对餐后血糖的升高起到降低的作用。随后通过改善肠道菌群的丰度和构成,影响肠道菌群进而对胰岛素信号传导通路、糖代谢和脂代谢进行调节,从而对糖尿病造成的一系列损伤进行修复。综上,乳酸菌能够通过改善胰岛素抵抗,调节胰岛素信号转导、脂质代谢、糖代谢,恢复肠道菌群对糖尿病造成的损伤进行改善。
在针对乳酸菌降血糖的研究中,主要集中在通过肠道菌群的重建和丰富菌群数量以调节其他代谢途径来治疗糖尿病引起的并发症,并对其具体机理进行探究。乳酸菌作为一种天然产物,其毒副作用小,效果好。未来作为糖尿病的辅助治疗药物或者潜在开发药物有着极大的前景。但目前食源性乳酸菌的临床试验还很少,糖尿病人机体防御力很弱,而乳酸菌摄入可能引发感染。因此对可食用乳酸菌的筛选是未来研究的重要方向。另外很多乳酸菌并不适宜在肠道内定植,因此通过生物工程的技术对乳酸菌进行改造也是未来乳酸菌研究新的思路和方向。
-
![]()
图 1 抑制α-葡萄糖苷酶降低血糖原理
Figure 1. Principle of inhibiting α-glucosidase to lower blood sugar
-
[1] Diabetes atlas—8th edition[M/OL]. Inter National Diabetes Federation, 2017. http://www.Diabetesatlas.org/resources/2017-atlas.
[2] Holman N, Young B, Gadsby R. Current prevalence of type 1 and type 2 dia-betes in adults and children in the UK[J]. Diabet Med,2015,32(9):1119−1120. doi: 10.1111/dme.12791
[3] Bruno G, Runzao C, Cavallo-Perin P, et al. Incidence of type 1 and type 2 diabetesin adults aged 30-49 years: The population-based registry in the province of Turin, Italy[J]. Diabetes Care,2005,28(11):2613−2619. doi: 10.2337/diacare.28.11.2613
[4] Cheng Y, Sibusiso L, Hou L, et al. Sargassum fusiforme fucoidan modifies the gut microbiota during alleviation of streptozotocin-induced hyperglycemia in mice[J]. International Journal of Biological Macromolecules,2019,131:1162−1170. doi: 10.1016/j.ijbiomac.2019.04.040
[5] Tank, Tesar C, Wiltonr, et al. Interaction of antidiabetic α-glucosidase inhibitors and gut bacteria α-glucosidase[J]. Protein Sci,2018,27(8):1498−1508. doi: 10.1002/pro.3444
[6] Duar R M, Lin X B, Zheng J, et al. Lifestyles in transition: evolution and natural history of the genus Lactobacillus[J]. FEMS Microbiol Rev,2017,41(Supp_1):S27−S48. doi: 10.1093/femsre/fux030
[7] O’Callaghan J, W O’Toole P. Lactobacillus: Host–microbe relationships[J]. Curr Top Microbiol Immunol,2013,358:119−54.
[8] Ghosh T, Beniwal A, Semwal A, et al. Mechanistic insights into probiotic properties of lactic acid bacteria associated with ethnic fermented dairy products[J]. Frontiers in Microbiology,2019,10:502. doi: 10.3389/fmicb.2019.00502
[9] Huang Y, Wang X, Wang J, et al. Lactobacillus plantarum strains as potential probiotic cultures with cholesterol-lowering activity[J]. Journal of Dairy Science,2013,96(5):2746−2753. doi: 10.3168/jds.2012-6123
[10] Huang Y C, Wu B H, Chu Y L, et al. Effects of tempeh fermentation with Lactobacillus plantarum and Rhizopus oligosporus on streptozotocin-induced type II diabetes mellitus in rats[J]. Nutrients,2018,10(9):1143. doi: 10.3390/nu10091143
[11] Mekkes M C, Weenen T C, Brummer R J, et al. The development of probiotic treatment in obesity: A review[J]. Benef Microbes,2014,5(1):19−28. doi: 10.3920/BM2012.0069
[12] Yan F, Li N, Shi J, et al. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice[J]. Food Funct,2019,10(9):5804−5815. doi: 10.1039/C9FO01062A
[13] 缪子敬, 李志万, 周学海, 等. α-葡萄糖苷酶抑制剂的研究进展[J]. 广东化工,2019,46(17):96−97, 101. doi: 10.3969/j.issn.1007-1865.2019.17.042 [14] Li X, Wang N, Yin B, et al. Effects of Lactobacillus plantarum CCFM0236 on hyperglycaemia and insulin resistance in high-fat and streptozotocin-induced type 2 diabetic mice[J]. J Appl Microbiol,2016,121(6):1727−1736. doi: 10.1111/jam.13276
[15] 党芳芳. 副干酪乳杆菌对糖尿病的改善作用及其机制研究[D]. 黑龙江: 东北农业大学, 2018. [16] Hansen J B, Arkhammar P O, Bodvarsdottir T B, et al. Inhibition of insulin secretion as a new drug target in the treatment of metabolic disorders[J]. Curr Med Chem,2004,11(12):1595−1615. doi: 10.2174/0929867043365026
[17] Li X, Wang E, Yin B, et al. Effects of Lactobacillus casei CCFM419 on insulin resistance and gut microbiota in type 2 diabetic mice[J]. Benef Microbes,2017,8(3):421−432. doi: 10.3920/BM2016.0167
[18] Korkmaz O A, Sumlu E, Koca H B, et al. Effects of Lactobacillus plantarum and Lactobacillus helveticus on renal insulin signaling, inflammatory markers, and glucose transporters in high-fructose-fed rats[J]. Medicina (Kaunas),2019,55(5):207. doi: 10.3390/medicina55050207
[19] Sumlu E, Bostancı A, Sadi G, et al. Lactobacillus plantarum improves lipogenesis and IRS-1/AKT/eNOS signalling pathway in the liver of high-fructose-fed rats[J]. Arch Physiol Biochem,2020:1−9.
[20] Fan C Y, Wang M X, Ge C X, et al. Betaine supplementation protects against high-fructose-induced renal injury in rats[J]. Nutr Biochem,2014,25(3):353−362. doi: 10.1016/j.jnutbio.2013.11.010
[21] Nizar J M, Shepard B D, Vo V T, et al. Renal tubule insulin receptor modestly promotes elevated blood pressure and markedly stimulates glucose reabsorption[J]. JCI Insight,2018,3(16):95−107.
[22] Rayasam G V, Tulasi V K, Sodhi R, et al. Glycogen synthase kinase 3: More than a namesake[J]. Br J Pharmacol,2009,156(6):885−898. doi: 10.1111/j.1476-5381.2008.00085.x
[23] Zarfeshani A, Khaza'ai H, Mohd Ali R, et al. Effect of Lactobacillus casei on the production of pro-inflammatory markers in streptozotocin-induced diabetic rats[J]. Probiotics Antimicrob Proteins,2011,3(3-4):168−174. doi: 10.1007/s12602-011-9080-9
[24] Yadav R, Dey D K, Vij R, et al. Evaluation of anti-diabetic attributes of Lactobacillus rhamnosus MTCC: 5957, Lactobacillus rhamnosus MTCC: 5897 and Lactobacillus fermentum MTCC: 5898 in streptozotocin induced diabetic rats[J]. Microb Pathog,2018,125:454−462. doi: 10.1016/j.micpath.2018.10.015
[25] Shih C C, Shlau M T, Lin C H, et al. Momordica charantia ameliorates insulin resistance and dyslipidemia with altered hepatic glucose production and fatty acid synthesis and AMPK phosphorylation in high-fat-fed mice[J]. Phytother Res,2014,28(3):363−371. doi: 10.1002/ptr.5003
[26] Taylor R, Al-Mrabeh A, Zhyzhneuskaya S, et al. Remission of human type 2 diabetes requires decrease in liver and pancreas fat content but is dependent upon capacity for β cell recovery[J]. Cell Metab,2018,28(4):667. doi: 10.1016/j.cmet.2018.08.010
[27] Liu D, Huang Y, Bu D, et al. Sulfur dioxide inhibits vascular smooth muscle cell proliferation via suppressing the Erk/MAP kinase pathway mediated by cAMP/PKA signaling[J]. Cell Death Dis,2014,5(5):e1251. doi: 10.1038/cddis.2014.229
[28] Hsieh F C, Lee C L, Chai C Y, et al. Oral administration of Lactobacillus reuteri GMNL-263 improves insulin resistance and ameliorates hepatic steatosis in high fructose-fed rats[J]. Nutr Metab (Lond),2013,10(1):35. doi: 10.1186/1743-7075-10-35
[29] Liu C, Shen Y J, Tu Q B, et al. Pedunculoside, a novel triterpene saponin extracted from Ilex rotunda, ameliorates high-fat diet induced hyperlipidemia in rats[J]. Biomed Pharmacother,2018,101:608−616. doi: 10.1016/j.biopha.2018.02.131
[30] 路晓杰, 刘久茜, 曹永国, 等. 普洱熟茶提取物对实验性非酒精性脂肪肝鼠脂代谢指标及肠道菌群的调节作用[J]. 中国兽医学报,2018,38(4):751−758. [31] Xie N, Cui Y, Yin Y N, et al. Effects of two Lactobacillus strains on lipid metabolism and intestinal microflora in rats fed a high-cholesterol diet[J]. BMC Complement Altern Med,2011,11:53. doi: 10.1186/1472-6882-11-53
[32] Qian Y, Li M, Wang W, et al. Effects of Lactobacillus casei YBJ02 on lipid metabolism in hyperlipidemic mice[J]. J Food Sci,2019,84(12):3793−3803. doi: 10.1111/1750-3841.14787
[33] Zhao Z, Wang C, Zhang L, et al. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway[J]. Appl Microbiol Biotechnol,2019,103(14):5843−5850. doi: 10.1007/s00253-019-09703-4
[34] Park D Y, Ahn Y T, Huh C S, et al. Dual probiotic strains suppress high fructose-induced metabolic syndrome[J]. World J Gastroenterol,2013,19(2):274−283. doi: 10.3748/wjg.v19.i2.274
[35] Horie M, Miura T, Hirakata S, et al. Comparative analysis of the intestinal flora in type 2 diabetes and nondiabetic mice[J]. Exp Anim,2017,66(4):405−416. doi: 10.1538/expanim.17-0021
[36] Vijay-Kumar M, Aitken J D, Carvalho F A, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5[J]. Science,2010,328(5975):228−231. doi: 10.1126/science.1179721
[37] Lin H V, Frassetto A, Kowalik E J Jr, et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms[J]. PLoS One,2012,7(4):e35240. doi: 10.1371/journal.pone.0035240
[38] Wang G, Li X, Zhao J, Zhang H, et al. Lactobacillus casei CCFM419 attenuates type 2 diabetes via a gut microbiota dependent mechanism[J]. Food Funct,2017,8(9):3155−3164. doi: 10.1039/C7FO00593H
[39] Li S, Qi C, Zhu H, et al. Lactobacillus reuteri improves gut barrier function and affects diurnal variation of the gut microbiota in mice fed a high-fat diet[J]. Food Funct,2019,10(8):4705−4715. doi: 10.1039/C9FO00417C
[40] Balakumar M, Prabhu D, Sathishkumar C, et al. Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice[J]. Eur J Nutr,2018,57(1):279−295. doi: 10.1007/s00394-016-1317-7
[41] Kim K A, Gu W, Lee I A, et al. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway[J]. PLoS One,2012,7(10):47713−47723. doi: 10.1371/journal.pone.0047713
[42] Toye A A, Dumas M E, Blancher C, et al. Subtle metabolic and liver gene transcriptional changes underlie diet-induced fatty liver susceptibility in insulin-resistant mice[J]. Diabetologia,2007,50(9):1867−1879. doi: 10.1007/s00125-007-0738-5
[43] Dumas M E. Barton R H, Toye A, et al. Metabolic profiling reveals a contribution of gut mierobiota to fatty liver phenotype in insulin-resistant mice[J]. Proc Natl Acad Sci USA,2006,103:12511−12516. doi: 10.1073/pnas.0601056103
 下载:
下载:
 下载:
下载:

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
