


Research Progress on the Regulation of Bacteriocin Synthesis by Environmental Stress in Lactobacillus plantarum
-
摘要: 植物乳杆菌作为具有重要经济价值的乳酸菌被广泛应用于食品发酵与保鲜领域,由于其代谢过程中会产生具有广谱抑菌特性、对热稳定且易被蛋白酶水解的细菌素,因此有作为天然食品生物防腐剂的较大应用潜力。研究表明,在发酵过程中菌体的生长和细菌素的合成受多种环境因素如盐胁迫、酸胁迫、氧胁迫及低高温胁迫的影响,但目前环境因素调节信号分子产生以及调控相关基因合成细菌素的具体机制仍然有待研究,另一方面,通用的调控通路还未被发现。因此,本文介绍了植物乳杆菌抵御胁迫的反应机制并详细阐述了环境胁迫下与细菌素合成密切相关的调控基因和重要调控蛋白,为食品发酵加工过程中合理控制发酵条件,促进细菌素合成从而延长食品货架期提供理论依据。Abstract: As a species of lactic acid bacteria with important economic value, Lactobacillus plantarum has been widely used in the field of food fermentation and preservation. Because it can produce bacteriocins with broad-spectrum antibacterial properties, thermal stability and easy to be hydrolyzed by proteases in the process of metabolism, Lactobacillus plantarum has great potential to be used as a natural food biological preservative. Studies have shown that the growth of bacteria and the synthesis of bacteriocin during fermentation are affected by various environmental factors, such as salt stress, acid stress, oxygen stress and low or high temperature stress. However, how environmental factors regulate the generation of signal molecules and the specific mechanism of environmental factors regulating the synthesis of bacteriocin by related genes need to be studied. On the other hand, general regulatory pathways have not been discovered. Therefore, this article reviews the response mechanism of Lactobacillus plantarum resisting stress. In addition, the regulatory genes and important regulatory proteins that are closely related to bacteriocin synthesis under environmental stress were introduced in detail. The above review would provide a theoretical basis for rationally controlling the fermentation conditions during food fermentation and promoting the synthesis of bacteriocin to extend the shelf life of food.
-
Keywords:
- environmental stress /
- bacteriocin /
- Lactobacillus plantarum /
- regulatory gene /
- regulatory protein
-
植物乳杆菌(Lactobacillus plantarum)是食品发酵技术中使用最为广泛的乳杆菌之一,已被证实有能力在人类和其他哺乳动物的肠道中定植[1],并具有良好的代谢和耐酸能力[2]。植物乳杆菌具有调节人体免疫[3]、抵制胃肠道致病菌感染的功效[4],同时其对肠上皮细胞具有较好的黏附能力,在强酸和高浓度胆盐下仍具有较高存活率[5],且其代谢过程中会产生细菌素等无毒副作用和耐药性的次级代谢产物[6]。细菌素是由核糖体合成并分泌至胞外介质的具备生物活性的多肽,当其达到一定浓度时可抑制亲缘关系相近的致病菌或腐败菌的生长[7],它们对热稳定且易被蛋白酶水解,因此细菌素作为生物防腐剂广泛应用于各种发酵食品[8]。
植物乳杆菌可以在食品基质中的热、冷、酸、盐等应激条件下存活,这表明它对于复杂恶劣环境已经形成了耐受性和抗性机制。国内外学者对植物乳杆菌在环境胁迫下的应激机制主要从分子机制和蛋白质组学角度进行研究,尽管一些应激基因已经在这个物种中被鉴定出来[9],但是对其转录调控机制仍然知之甚少。有研究表明环境胁迫可以正向调控细菌素的合成,Leroy等[10]研究了不同环境胁迫对Enterococcus faecium RZSC5产生细菌素的调控作用,研究发现中等盐浓度能够促进细菌素的分泌。Hurtado等[11]研究了Lactobacillus pentosus B96在盐胁迫下细菌素编码基因表达的变化,研究表明4%和6%浓度的盐胁迫可以促进细菌素的生物合成,plnE和plnN基因的表达受胁迫影响显著上调,但其并未深入研究盐胁迫如何调控细菌素的合成。乳酸菌细菌素虽然具备无毒、高效抑菌等优点,但产量较低,在工业化批量生产中大大受限。
目前利用基因工程技术提高细菌素产量虽理论上可行,但普遍存在细菌素活性较低及安全问题。而环境胁迫对细菌素的调控是基于转录水平上的调节,并不涉及安全性问题。由于环境胁迫调控细菌素合成的机制较为复杂,调控因子及调控通路尚不明确,通过施加环境压力提升菌体细菌素合成量的方法依然难以投入到实际应用中。因此完善关于环境胁迫对细菌素合成的分子调控机制的研究,对促进生物保护发酵剂的发展有着重要意义。
1. 植物乳杆菌细菌素合成的分子调控机制
1.1 Al-2介导的群体感应系统
群体感应(quorum sensing,QS)是指细菌感应外界环境刺激,分泌自诱导物或自诱导多肽作为信号分子,当其浓度达到一定阈值时会启动菌体某些功能性基因的表达进而影响菌体生命活动,如调控细菌素的合成及分泌[12]。Saucier等[13]在Carnobacterium piscicola LV17中首次发现乳酸菌存在群体感应现象。少数Ⅰ类和多数Ⅱ类细菌素的生物合成受QS系统调控。调节Ⅱ类细菌素产生的QS系统由自诱导肽(auto-inducing peptide,AIP)、监测环境的膜定位组氨酸蛋白激酶(histidine protein kinase,HPK)以及细胞质感应调节蛋白(response regulator,RR)等3种调控蛋白组成,被称为三组分系统,其中HPK和RR也被称为双组份系统(two component system,TCS)[14]。Al-2以具备生物活性的信号分子身份参与QS信息交流进程,从分子结构层面来看,Al-2是一种呋喃酰硼酸二酯类化合物[15]。它由luxS基因编码的关键酶催化S-核糖高半胱氨酸(SRH)生成,其中SRH是通过pfs基因编码合成酶催化S-腺苷高半胱氨酸(SAH)而形成的。植物乳杆菌与其他诱导菌共培养时细菌素也会被诱导进而杀死或抑制其它微生物。Di等[16]研究发现共培养后L. plantarum DC400细菌素的合成量显著上升,luxS基因表达量也较纯培养显著提升,由此推断luxS介导的群体感应系统间接调控植物乳杆菌细菌素的合成。Man等[17]研究发现L. plantarum KLDS1.0391与瑞士乳杆菌KLDS1.9207共培养时,编码细菌素的pln基因簇相关基因的表达显著上调。与此同时,Ruiz-Barba等[18]研究发现,L. plantarum NC8与戊糖片球菌FBB63等革兰氏阳性菌共培养时,L. plantarum NC8细菌素调节基因转录效率有所上升。综上表明,细菌素产量的提高可能是微生物的存在刺激生产者生长的结果,也可能是自诱导物Al-2产量增加的结果。
1.2 植物乳杆菌pln基因簇
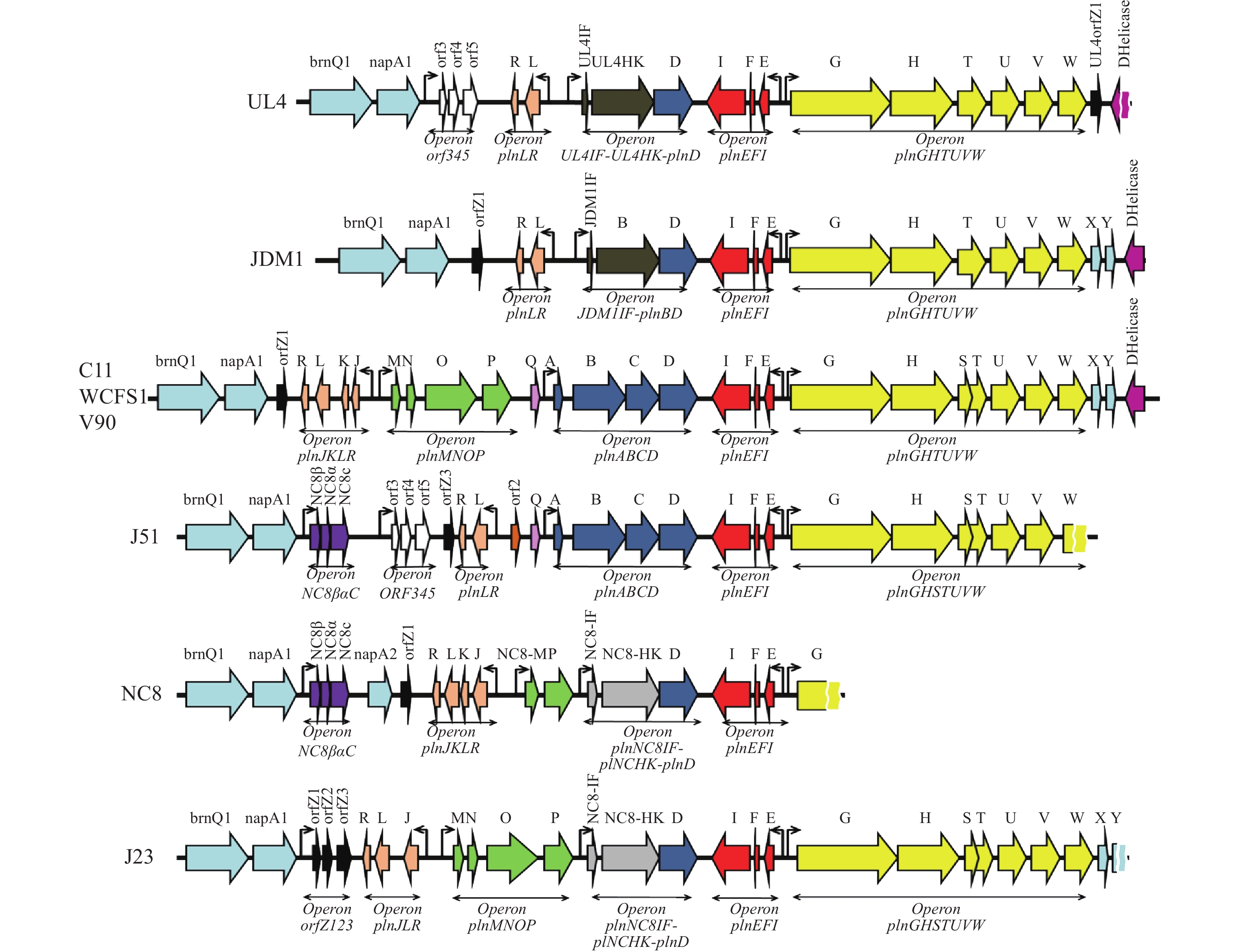
植物乳杆菌细菌素调控系统除上述提到的Al-2介导的群体感应系统以外,还受基因组中pln基因簇的调控。Saenz等[19]证实了L. plantarum C11、WCFS1、J51、YM-4-3中均含有5个诱导操纵子:分别是plnABCD、plnJKLR、plnEFI、plnGHSTUVW、plnMNOP。各诱导操纵子的具体功能如下:负责编码群体感应系统并可启动其他操纵子转录起始的调节操纵子plnABCD,其中plnA编码自诱导肽(AIP),plnB编码可跨膜的组氨酸激酶(HPK),plnC和plnD编码两种功能相反的反应调节因子[20],plnC编码的蛋白激活基因簇转录,plnD编码的蛋白则起抑制作用;编码细菌素结构基因的plnJKLR和plnEFI,其中PlnI及PlnL为abi基因编码免疫蛋白;编码ABC转运系统并参与细菌素分泌运输的运输操纵子plnGHSTUVW;plnMNOP等功能还未可知。Diep等[21]认为plnEFI和plnJKLR操纵子编码双肽细菌素、PlnEF和PlnJK及其相应的免疫蛋白。plnGHSTUVWXY操纵子编码ABC转运系统的蛋白质,该系统负责分泌并处理细菌素前体。2003年,Diep等[22]进一步验证了PlnA肽诱导上述五个操纵子的转录的同时,自身也是一种抑菌物质。Maldonado-Barragan等[23]通过基因敲除实验验证了L. plantarum WCFS1中的plnABCD以及L. plantarum NC8中的plnNC8IF-HK-D对细菌素的合成至关重要。Tai等[24]对L. plantarum WCFS1、C11、V90、J23、J51、NC8、JDM1、UL4的pln基因簇进行了总结(图1)。
![]()
2. 植物乳杆菌抵御胁迫的反应机制
2.1 植物乳杆菌面临的环境胁迫
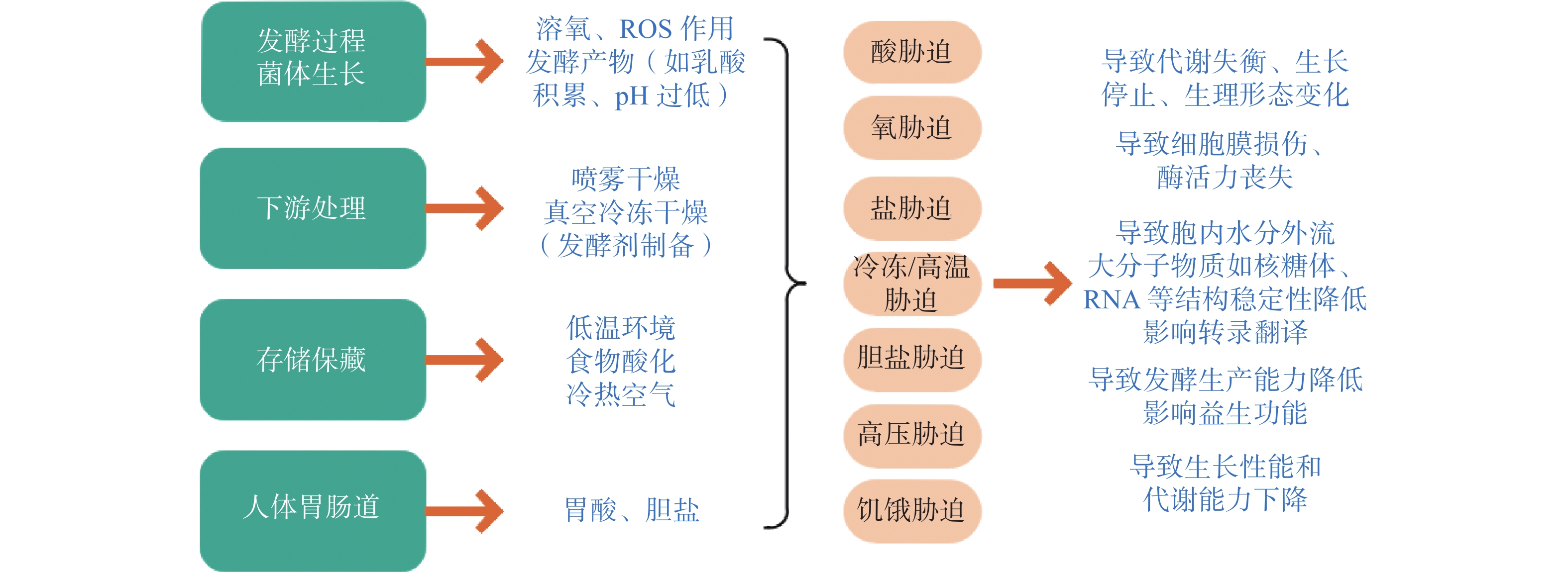
利用植物乳杆菌产细菌素的特性提高食品安全性是食品防腐保鲜的一种手段,但无论是在加工运输中还是摄入体内发挥其益生效应均会面临诸多胁迫(图2)。在菌体生长、存储保藏、经过人体肠胃道等过程中,菌体自身会受酸、冷冻、高温、盐、饥饿等胁迫作用而损伤甚至死亡。例如人体胃肠道的低pH环境或食品加工基质中遇到的酸胁迫,会导致胞内质子积累同时降低跨膜推动力,进而降低对酸敏感的酶活性,也会损害蛋白质和DNA等生物大分子;作为发酵剂在加工贮存中遇到的低温环境不仅会减弱细胞膜流动性,还会影响基因表达使菌体产生冷休克反应,相反,环境温度过高又会致使蛋白降解;细菌代谢过程产生的活性氧(ROS)、超氧阴离子(O2−)、过氧化氢(H2O2)以及过氧化自由基(O·)等物质会使菌体细胞受损[25];细菌繁殖过程中,若营养物质得不到及时补充,便会引发营养胁迫现象,导致代谢能力大大降低,如细菌素等代谢产物合成量降低。
![]() 图 2 植物乳杆菌面临的环境胁迫Figure 2. Environmental stress encountered by Lactobacillus Plantarum
图 2 植物乳杆菌面临的环境胁迫Figure 2. Environmental stress encountered by Lactobacillus Plantarum2.2 植物乳杆菌抵御盐胁迫的调控机制
植物乳杆菌抵御盐胁迫通常依靠相容性溶质调控系统、胞内离子平衡、热休克蛋白调控系统及其他途径关键酶调控系统。在高渗透压条件下,植物乳杆菌主要通过从细胞外部吸收甘氨酸甜菜碱(glycine betaine,GB)等相容性溶质来抵御盐胁迫[26]。其中L. plantarum主要采取QacT系统转运相似相容类物质[27],有研究指出L. plantarum ATCC14917依靠QacT系统转运甜菜碱、肉碱和脯氨酸[28],而L. plantarum WCFS1拥有两个相容性溶质转运系统,即ChoQS、OpuABCD[29]。植物乳杆菌还可以利用质膜上的Na+/H+逆向转运蛋白将胞内Na+外排,进而维持胞内离子平衡[30]。此外,有研究报道指出L. plantarum ST-III中存在编码Na+/H+逆向转运蛋白的基因[31]。近期还有学者发现,L. plantarum ST-III的质粒上包含一个由kdpABCDE操纵子编码的K+转运基因簇[32]。另一方面,当菌体外部渗透压升高时,编码普遍应激蛋白(GSPs)、热激蛋白(HSPs)和盐胁迫蛋白(SSPs)的基因表达量会显著上升。HSPs作为分子伴侣蛋白通常包括DnaK、GroEL、DnaJ和GroES,在蛋白折叠修复和降解中起到关键作用[33]。也有研究发现,胞内蛋白FtsH和细胞看家蛋白HtrA在盐胁迫下表达上调。糖酵解途径是植物乳杆菌生成乳酸的重要途径,有学者指出糖酵解关键酶基因pfk、fba、pgk、ldh的表达也会随着NaCl浓度的升高而上调。盐胁迫也可以诱导腺苷酸琥珀酸合酶PurA的合成,有研究发现参与嘌呤代谢的PurH蛋白在5 mol/L NaCl胁迫下表达上调。这些关键酶数量的增加可以使核苷酸积累用于DNA、RNA和ATP的合成,维持菌体正常生长代谢。
2.3 植物乳杆菌抵御酸胁迫的调控机制
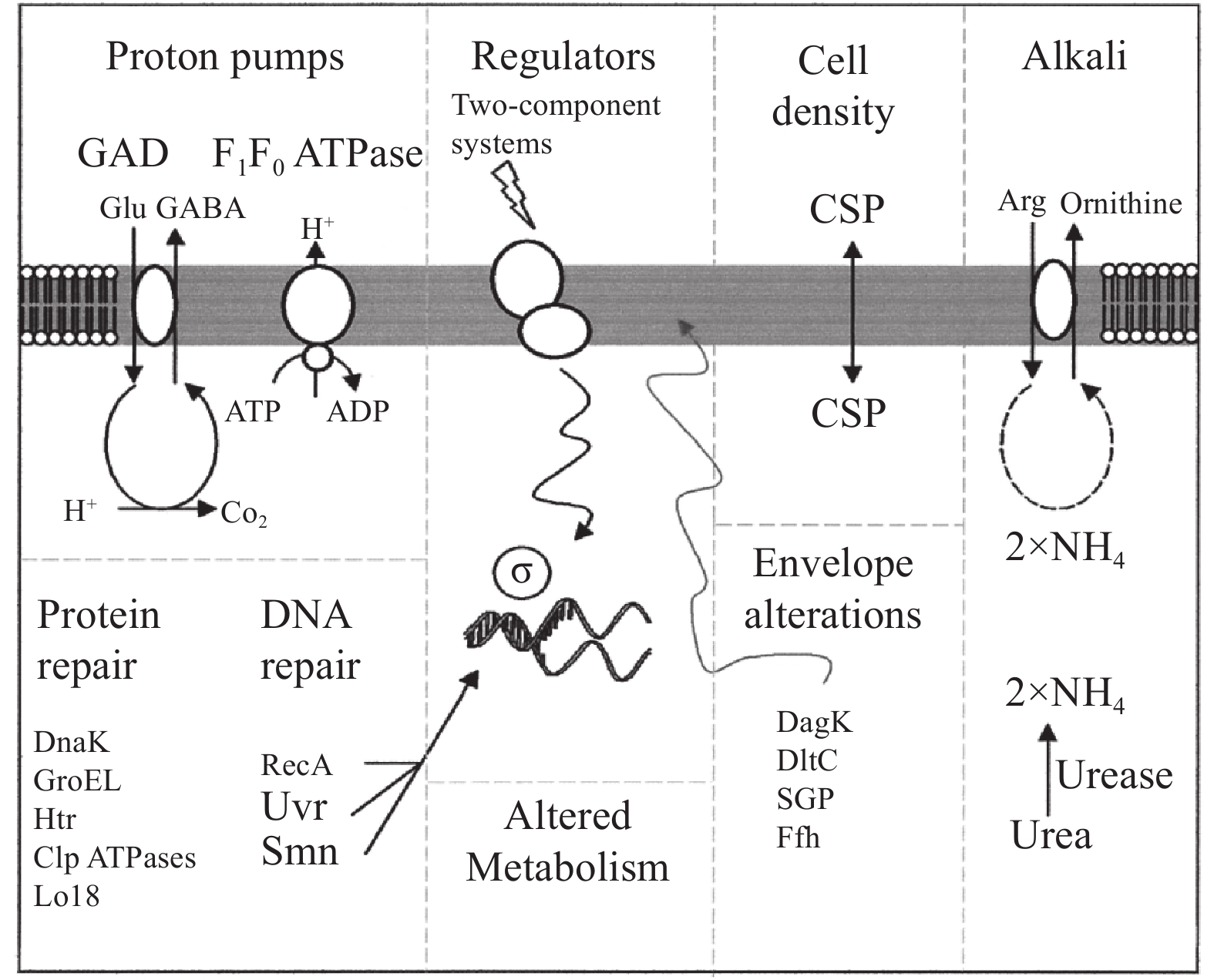
植物乳杆菌可以通过调控胞内pH动态平衡、改变细胞膜生理功能以及分泌抗胁迫应激蛋白等方式来应对发酵过程中所产有机酸带来的酸胁迫。胞内pH的动态平衡对于L. plantarum抵御酸胁迫至关重要,乳酸菌能够通过H+-ATPase、谷氨酸脱氢酶体系,精氨酸脱羧酶途径等机制维持胞内pH的动态平衡(图3)[34]。当胞内pH处于菌体能耐受的范围外时,F0F1-ATPase聚合体会消耗胞内ATP,将胞内水解产物H+泵出至胞外,以维持胞内pH的稳定。此外,菌体的精氨酸脱氨酶体系可以通过催化精氨酸产生NH3,随后胞内H+与产物NH3结合,从而使pH升高至正常范围,此过程产生的ATP有助于F0F1-ATPase泵出质子。细胞膜的结构成分在抵御酸胁迫的过程中也起到关键作用,如调整膜脂肪酸中的长链脂肪酸和单不饱和脂肪酸的比例。有研究尝试阐明植物乳杆菌的耐酸机制,Liu等[35]认为植物乳杆菌SA-680包含一个耐酸基因mdt,其序列与多种植物乳杆菌膜转运蛋白基因编码区序列很接近,猜测基因mdt可能与质子流动有关。
2.4 植物乳杆菌抵御冷热胁迫的调控机制
植物乳杆菌作为冷冻发酵剂在储存过程中、干酪成熟阶段及冷藏产品的低温发酵阶段会引发大量的冷诱导蛋白(Cold Induced Proteins,CIP)表达。Derzelle等[36]从受到冷胁迫的植物乳杆菌Lp80和C3.8中分别克隆出cspL和cspP两种冷休克蛋白基因。此外,工业生产中喷雾干燥、巴斯德灭菌等高温过程带来的热应激会导致DnaK、DnaJ、HrcA、GroES、GroEL、Hsp84、Hsp85、Hsp100、C1p、HtrA和FtsH等热休克蛋白和蛋白酶表达上调。Castaldo等[37]研究发现,L. plantarum LM3-2含有以cis启动的CIRCE序列为特征的DnaK和GroESL热激操纵子。刘倩颖[38]通过KEGG通路富集结果分析出L. plantarum LIP-1在冷胁迫下嘌呤代谢通路最显著,冷应激蛋白CspC的表达显著上调。
2.5 植物乳杆菌抵御氧胁迫的调控机制
植物乳杆菌的呼吸代谢产物活性氧(ROS),包括超氧阴离子自由基(O2−),过氧化氢(H2O2)和羟自由基(OH-)等,这类物质会打破胞内的氧化还原平衡进而促使ROS浓度升高进而造成氧胁迫。乳酸菌抵御氧胁迫多依赖于SOD、CAT和GSH-PX等ROS解毒酶类。Eric等[39]发现在L. plantarum中,两个谷胱甘肽还原酶同源物可以有效降低由胆盐胁迫引起的氧化损伤。与此同时,过氧化物响应调控因子PerR、氧气响应转录调节子FNR以及Rex类转录因子在抵抗氧化胁迫和生物膜形成方面也有重要调控作用。另一方面,FlpA和FlpB通过上调Zn2+的摄取来增加菌体对胞内巯基蛋白的保护从而提高菌体氧化胁迫抗性。此外,有研究为阐明L. plantarum CAUH2的抗氧化胁迫机制,采用转录组学进行分析,结果表明转录因子GopR具有抗氧化功能。
3. 环境胁迫对植物乳杆菌细菌素合成量的影响
不同植物乳杆菌菌株由于自身的差异性,面对不同胁迫时其细菌素的合成能力存在较大差异。食品基质中的低温、低pH和高NaCl等胁迫因素会影响乳酸菌的生长和细菌素的合成。Engelhardt等[40]研究结果指出L. plantarum ESB202与P. acidilactici HA6111-2在7.5% NaCl浓度下分别培养,二者细菌素抑菌活性均被显著抑制。而Lim等[41]研究发现适宜的NaCl浓度(1%或3%)使L. plantarum KC21细菌素的合成量翻倍,抑菌活性升至12800 BU/mL,当NaCl浓度大于5%时会抑制菌株的生长和细菌素的合成。Leal-Sanchez等[42]同样发现低浓度的NaCl胁迫(2.3%~2.5%)也会促进L. plantarum LPCO10细菌素的合成。此外,Vazquez等[43]发现外源添加1.5 g/L的半胱氨酸或色氨酸对Nisin的合成有促进作用。Yi等[44]以Glu、Gly、Cys、Tyr和Ala为研究对象,其结果表明只有Cys和Gly对细菌素的合成有诱导作用,其余氨基酸作用不显著。但上述氨基酸是通过影响菌体生长进而影响细菌素的产量,还是作为前体物质直接参与细菌素的合成或是作为中间调控因子还有待深入研究。另一方面,Parlindungan等[45]发现L. plantarum B21受到营养胁迫如葡萄糖胁迫时,生长速度明显降低,且不产生酸,但细菌素的活性仍能被检测。与此相反,受吐温80的胁迫时,尽管生长速度快、产酸量大,但细菌素活性不明显。上述结果都仅为表观研究,没有针对内在机制进行深入探讨。
4. 环境胁迫调控细菌素合成的分子机制
4.1 细胞分泌途径中重要调控基因及蛋白
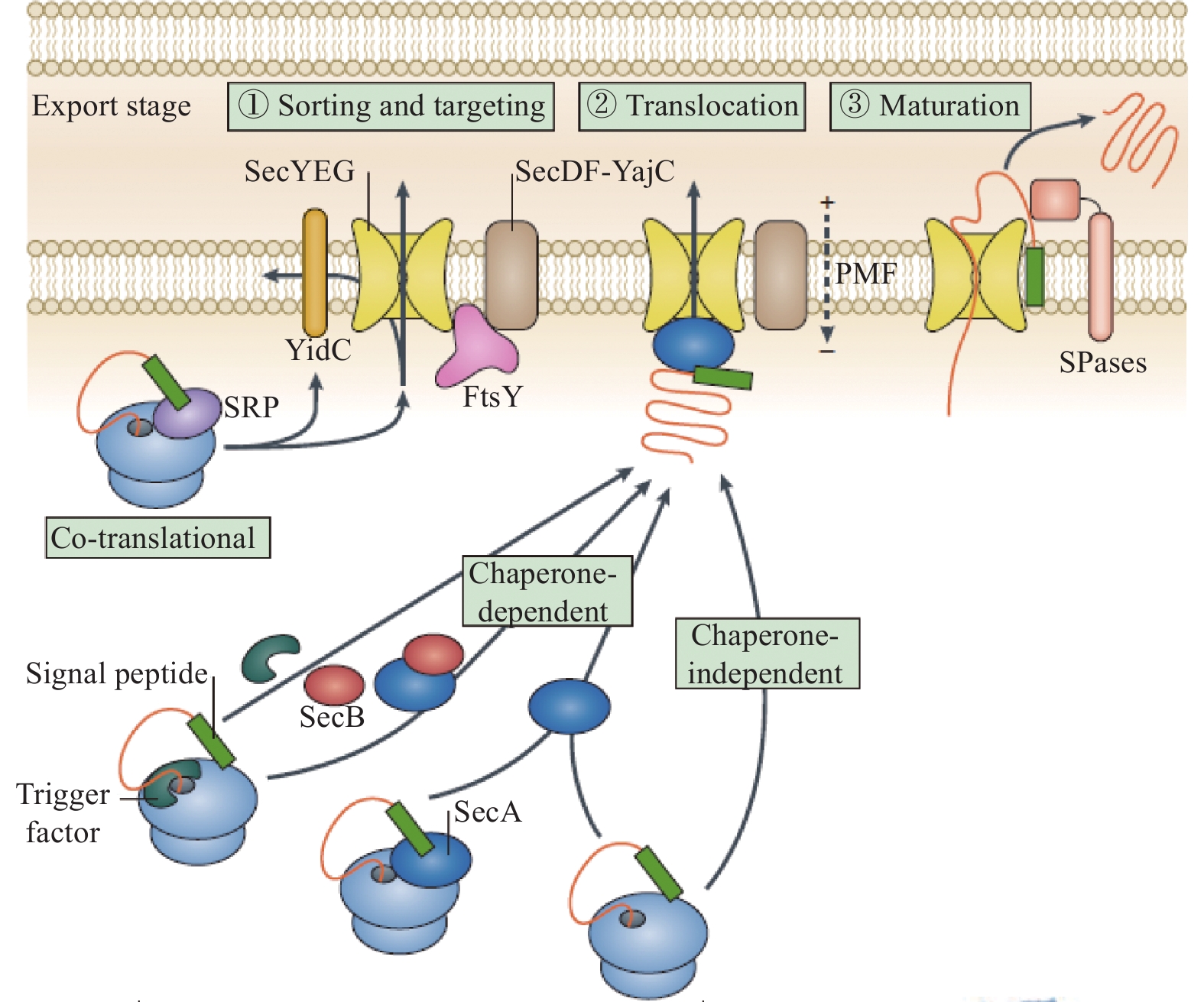
生物体面对压力变化时通常会诱导大量基因表达并合成应激蛋白质,进而适应不良环境。也有报道指出,菌体面临环境胁迫会激活多种代谢途径[46]。细菌素作为一种分泌蛋白,在其运输分泌过程中ABC转运蛋白作为一种载体起主要作用。同时,由于细菌素自身结构及底物信号序列的不同,其分泌转运途径可分为NisT型、SunT型、Sec型、SRP型途径以及其他转运蛋白途径[47](图4)。其中Sec型分泌途径是细菌蛋白分泌到胞外的常见方式之一,有研究报道指出,SecY、SecE和SecG三种调控蛋白共同构成细菌蛋白异位复合体,形成蛋白转运通道,该分泌途径中SecA蛋白是分泌蛋白跨内膜转运的关键蛋白。此外,有研究表明,当菌体面临氧化应激时,分泌调控蛋白SecA的C末端特异性结合一种胞质蛋白MrgA,据报道该蛋白与氧化应激有关[48]。除此之外,SRP型蛋白分泌途径也较为常见,大多数膜蛋白和分泌蛋白在其靶向定位过程中都会受信号识别颗粒(signal recognition particle,SRP)的严格调控。目前针对乳酸菌SRP途径的研究主要集中在信号识别颗粒Ffh、受体FtsY的结构与功能。Froderberg等[49]研究发现蛋白转运酶YidC在细菌素分泌过程及Sec易位过程中均起重要作用,信号识别颗粒Ffh可与细菌素形成复合体,再与受体FtsY结合并定位于细胞膜。另一方面,Neumann等[50]认为信号识别颗粒SRP和分泌调控蛋白SecA在分泌蛋白跨内膜转运过程中起协同作用,二者可协助细菌蛋白的分泌。还有研究报道,当菌体面临盐胁迫时,busR基因编码的调控子会刺激BusA转运蛋白的表达,进而影响细菌素的分泌[51]。综上所述,胁迫下细菌分泌系统相关基因的表达上调使得蛋白共翻译转运速率提升,分泌蛋白的数量增加,由此推测胁迫通过影响细胞膜分泌系统进而影响细菌素的合成。
4.2 氨基酸代谢途径中重要调控基因及蛋白
天冬氨酸及谷氨酸代谢途径是合成缬氨酸、组氨酸、脯氨酸、赖氨酸以及色氨酸等多种氨基酸的有效途径,这些氨基酸可以进一步被生物体利用合成细菌素等相关蛋白。宋雪飞等[53]的研究结果显示基因asnA、hisA、hisC、metE在15 g/L浓度的NaCl胁迫下表达均显著下调,其中asnA、hisA、metE分别是参与天冬氨酸代谢、组氨酸代谢和半胱氨酸代谢的关键基因。同时,有研究报道指出,盐胁迫下L. plantarum WCFS1中除了参与丝氨酸代谢途径的基因表达下调,编码氨基酸转运蛋白的基因表达也显著下调。细菌素本质也是一种多肽,其合成过程受上述氨基酸代谢的影响,这也在一定程度上解释了胁迫下细菌素合成量发生变化的原因。
另一方面,由argG和argH基因编码的ArgG和ArgH蛋白在天冬氨酸转化为精氨酸过程中起到关键的催化作用,张明阳[54]发现酸胁迫环境下过量表达上述两种蛋白会提高氨基酸代谢基因如精氨酸代谢基因arcA、arcB和argR的转录效率。其中arcA、arcB、argR、argG和argH等基因编码的蛋白能够使精氨酸分解并产生氨和ATP,由此推测上述基因可间接为细菌素的合成提供原料及能量,利于菌体抵抗酸胁迫[55]。同时,有研究发现随着外界渗透压升高,编码氨基肽酶PepX的基因pepX表达显著上调,该基因是调节蛋白质水解系统的关键基因,推断其与细菌素的水解及转运有关。
4.3 脂肪酸代谢途径中重要调控基因及蛋白
乳酸菌面临环境胁迫时,脂肪酸代谢途径中的酰基载体蛋白聚合物FabZ2、FabG2、FabH作为主导酶发挥优势作用,菌体生长速率加快,进而导致细菌素合成量显著升高。相反,Heunis等[56]发现酸胁迫条件下L. plantarum 423胞内FabD蛋白质的表达呈下降趋势,推断由于脂肪酸生物合成量减少进而抑制菌体自身生长,最终细菌素合成量随菌体生长能力减弱而降低。此外,还有研究表明基因accA、accB、accC、fabD及fabZ也参与脂肪酸生物合成途径,其中基因accA、accB及accC负责编码乙酰辅酶A羧化酶,基因fabD负责编码丙二酰辅酶A[57]。王茜茜等[58]基于iTRAQ技术发现盐胁迫会导致细胞膜合成相关基因fabG和fabZ表达上调,促进支链脂肪酸的合成从而提高细胞膜的流动性,进而利于细菌素的翻译和分泌。
4.4 核苷酸代谢途径中重要调控基因及蛋白
嘌呤核苷酸不仅作为DNA与RNA分子合成的主要底物,同时也参与ATP和GTP等物质的合成代谢,为细菌素合成及运输供给能量。嘌呤代谢通路的上游基因purL调控肌苷一磷酸(IMP)与焦磷酸硫胺素(TPP)的生物合成,二者对于细菌素及毒力因子的合成有一定影响。此外,基因sat及编码RNA聚合酶的基因rpoB过表达均会导致胞内ATP产量增多,为机体的生长代谢提供了更多的能量,有助于细菌素的合成与分泌。有学者基于转录组测序技术发现盐胁迫下L. plantarum WCFS1中编码嘌呤合成的基因purD、编码腺嘌呤琥珀酸合酶的基因purA、编码腺嘌呤脱氢酶的基因adeC、编码单磷酸鸟苷还原酶的基因guaC的表达均显著下调,直接导致嘌呤代谢过程中关键酶的活性减弱,间接导致细菌素产量下降[59]。Sun等[60]的研究结果与上述结果相似,差异蛋白结果显示酸胁迫条件下菌体中基因adk与基因purA的表达也有所下调。此外,Pang等[61]研究表明,胞内次黄嘌呤的浓度会随着基因purA转录水平的降低而增加,最终导致RNA和DNA合成量升高。由于细菌素作为一种分泌肽,RNA和DNA又作为其合成模板,因此推断出当菌体受胁迫时GuaC、PurH、PurA和PurB等蛋白质对细菌素有重要调控作用。综上所述,胁迫可以通过影响核苷酸代谢途径中能量的生成进而影响细菌素的合成。
4.5 其他重要调控基因及蛋白
核糖体蛋白、DNA修复蛋白、分子伴侣以及热休克蛋白作为全局调控蛋白,在乳酸菌胁迫响应中也发挥着重要作用。其中,分子伴侣蛋白如GroEL、GrpE、DnaK、DnaJ、Hsp、CspC等的主要作用是协助蛋白进行组装折叠、转运和降解,胁迫条件下这类蛋白质还能增加代谢相关蛋白如细菌素的稳定性[62]。1990年,Vanbogelen等[63]发现核糖体蛋白不仅具有促进蛋白质合成的功能,还能感受外界环境如热、冷、酸、渗透压等的变化并参与细菌素的翻译。2013年,陈卫等[64]发现当乳酸菌受酸、胆盐等环境胁迫时,热休克蛋白、冷休克蛋白和通用应激蛋白等通常被诱导表达。还有研究显示,大肠杆菌应对环境压力所采用的修复系统由全局转录诱导因子RecA和转录抑制因子LexA控制,自然状态下LexA蛋白结合在大肠杆菌素结构基因的启动子处,因此显著抑制细菌素合成[65]。然而当菌体受到紫外线等胁迫诱导后,RecA受到激活,促进阻遏蛋白LexA自身发生裂解,解除对细菌素结构基因的抑制作用,最终导致细菌素合成量增大。
5. 总结与展望
由于调控细菌素合成基因的多样性及其调控机制的复杂性,目前关于环境胁迫下植物乳杆菌细菌素合成机制的研究相对缺乏,仍有诸多问题有待解决,笔者认为主要集中在以下几个方面:第一,环境胁迫介导哪些基因的转录以及蛋白质的表达参与细菌素的合成;第二,是否存在相关调控蛋白与植物乳杆菌细菌素启动子调控区域有结合作用;第三,目前还没有找到通用的代谢通路用于解释环境胁迫下植物乳杆菌的应激机制;第四,应对不同环境胁迫时,群体感应系统相关的基因如luxS基因的转录水平、信号分子AI-2的活性及LuxS蛋白的稳定性如何变化还未可知。但随着深入研究,利用转录组和蛋白质组等多组学技术挖掘植物乳杆菌中重要功能基因,从转录水平、蛋白水平和代谢水平多层面研究不同胁迫下细菌素合成系统的表达差异是未来的研究方向。利用胁迫应答等外源压力提高植物乳杆菌细菌素产量的方法必将成为食品领域的研究热点,进而更好的发挥生物保护发酵剂的生防作用。
-
![]()
![]()
图 2 植物乳杆菌面临的环境胁迫
Figure 2. Environmental stress encountered by Lactobacillus Plantarum
-
[1] Butorac K, Banic M, Novak J, et al. The functional capacity of plantaricin-producing Lactobacillus plantarum SF9C and S-layer-carrying Lactobacillus brevis SF9B to withstand gastrointestinal transit[J]. Microbial Cell Factories,2020,19(1):106. doi: 10.1186/s12934-020-01365-6
[2] Chen P T, Hong Z S, Cheng C L, et al. Exploring fermentation strategies for enhanced lactic acid production with polyvinyl alcohol-immobilized Lactobacillus plantarum 23 using microalgae as feedstock[J]. Bioresource Technology,2020,308:123266. doi: 10.1016/j.biortech.2020.123266
[3] Hong Y F, Kim H, Kim H R, et al. Different immune regulatory potential of Lactobacillus plantarum and Lactobacillus sakei isolated from kimchi[J]. Journal of Microbiology and Biotechnology,2014,24(12):1629−1635. doi: 10.4014/jmb.1406.06062
[4] Liu J, Gu Z, Zhang H, et al. Preventive effects of Lactobacillus plantarum ST-III against Salmonella infection[J]. Lwt-Food Science and Technology,2019,105:200−205. doi: 10.1016/j.lwt.2019.02.043
[5] E Jingjing, Ma L, Chen Z, et al. Effects of buffer salts on the freeze-drying survival rate of Lactobacillus plantarum LIP-1 based on transcriptome and proteome analyses[J]. Food Chemistry,2020,326:126849. doi: 10.1016/j.foodchem.2020.126849
[6] Jin J, Jie L M, Zhang H, et al. Pediocin AcH is transcriptionally-regulated by a two-component system in Lactobacillus plantarum subs. plantarum[J]. Journal of Food Protection,2020,17(11):23−27.
[7] Lin T H, Pan T M. Characterization of an antimicrobial substance produced by Lactobacillus plantarum NTU 102[J]. Journal of Microbiology Immunology and Infection,2019,52(3):409−417. doi: 10.1016/j.jmii.2017.08.003
[8] Canak I, Markov K, Jakopovic Z, et al. Application of L. plantarum O1 producer of plantaricin NC8 (PLNC8) in biopreservation of aquatic food products[J]. Journal of Biotechnology,2017,256:S65−S65.
[9] Yao W, Yang L, Shao Z, et al. Identification of salt tolerance-related genes of Lactobacillus plantarum D31 and T9 strains by genomic analysis[J]. Annals of Microbiology,2020,70(1):3−9. doi: 10.1186/s13213-020-01549-w
[10] Leroy F, Vankrunkelsven S, De Greef J, et al. The stimulating effect of a harsh environment on the bacteriocin activity by Enterococcus faecium RZS C5 and dependency on the environmental stress factor used[J]. International Journal of Food Microbiology,2003,83(1):27−38. doi: 10.1016/S0168-1605(02)00316-1
[11] Hurtado A, Reguant C, Bordons A, et al. Expression of Lactobacillus pentosus B96 bacteriocin genes under saline stress[J]. Food Microbiology,2011,28(7):1339−1344. doi: 10.1016/j.fm.2011.06.004
[12] Maldonado-Barragan A, West S A. The cost and benefit of quorum sensing-controlled bacteriocin production in Lactobacillus plantarum[J]. Journal of Evolutionary Biology,2020,33(1):101−111. doi: 10.1111/jeb.13551
[13] Saucier L, Poon A, Stiles M E. Induction of bacteriocin in Carnobacterium piscicola LV17[J]. Journal of Applied Microbiology,2010,78(6):684−690.
[14] 崔艳华, 曲晓军, 马莺. 双组分系统分析预测共生机制[J]. 哈尔滨工业大学学报,2010,42(11):1798−1804. doi: 10.11918/j.issn.0367-6234.2010.11.026 [15] Medarametla P, Gatta V, Kajander T, et al. Structure-based virtual screening of LsrK kinase inhibitors to target quorum sensing[J]. Chemmedchem,2018,13(22):2400−2407. doi: 10.1002/cmdc.201800548
[16] Di Cagno R, De Angelis M, Coda R, et al. Molecular adaptation of sourdough Lactobacillus plantarum DC400 under co-cultivation with other lactobacilli[J]. Research in Microbiology,2009,160(5):358−366. doi: 10.1016/j.resmic.2009.04.006
[17] Man L L, Meng X C, Zhao R H, et al. The role of plNC8HK-plnD genes in bacteriocin production in Lactobacillus plantarum KLDS1.0391[J]. International Dairy Journal,2014,34(2):267−274. doi: 10.1016/j.idairyj.2013.08.009
[18] Ruiz-Barba J L, Caballero-Guerrero B, Maldonado-Barragán A, et al. Coculture with specific bacteria enhances survival of Lactobacillus plantarum NC8, an autoinducer-regulated bacteriocin producer, in olive fermentations[J]. Food Microbiology,2010,27(3):413−417. doi: 10.1016/j.fm.2009.10.002
[19] Saenz Y, Rojo-Bezares B, Navarro L, et al. Genetic diversity of the pln locus among oenological Lactobacillus plantarum strains[J]. International Journal of Food Microbiology,2009,134(3):176−183. doi: 10.1016/j.ijfoodmicro.2009.06.004
[20] Straume D, Johansen R F, Bjoras M, et al. DNA binding kinetics of two response regulators, PlnC and PlnD, from the bacteriocin regulon of Lactobacillus plantarum C11[J]. Bmc Biochemistry,2009,10(1):1−11. doi: 10.1186/1471-2091-10-1
[21] Diep D B, Johnsborg O, Risoen P A, et al. Evidence for dual functionality of the operon plnABCD in the regulation of bacteriocin production in Lactobacillus plantarum[J]. Molecular Microbiology,2010,41(3):633−644.
[22] Diep D B, Ronny M, Ola J, et al. Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln operons and by two antagonizing response regulators, the activity of which is enhanced upon phosphorylation[J]. Molecular Microbiology,2010,47(2):483−494.
[23] Maldonado-Barragan A, Ruiz-Barba J L, Jimenez-Diaz R. Knockout of three-component regulatory systems reveals that the apparently constitutive plantaricin-production phenotype shown by Lactobacillus plantarum on solid medium is regulated via quorum sensing[J]. International Journal of Food Microbiology,2009,130(1):35−42. doi: 10.1016/j.ijfoodmicro.2008.12.033
[24] Tai H F, Foo H L, Rahim R A, et al. Molecular characterisation of new organisation of plnEF and plw loci of bacteriocin genes harbour concomitantly in Lactobacillus plantarum I-UL4[J]. Microbial Cell Factories,2015,14:89. doi: 10.1186/s12934-015-0280-y
[25] Zhai Z, Yang Y, Wang H, et al. Global transcriptomic analysis of Lactobacillus plantarum CAUH2 in response to hydrogen peroxide stress[J]. Food Microbiology,2020,87:103389. doi: 10.1016/j.fm.2019.103389
[26] Wu R, Song X, Liu Q, et al. Gene expression of Lactobacillus plantarum FS5-5 in response to salt stress[J]. Annals of Microbiology,2016,66(3):1181−1188. doi: 10.1007/s13213-016-1199-1
[27] 林松洋, 郝利民, 刘鑫, 等. 乳酸菌耐盐分子机制研究进展[J]. 食品科学,2018,39(3):295−301. doi: 10.7506/spkx1002-6630-201803044 [28] Glaasker E, Heuberger E H M L, Konings W N, et al. Mechanism of osmotic activation of the quaternary ammonium compound transporter (QacT) of Lactobacillus plantarum[J]. Journal of Bacteriology,1998,180(21):5540. doi: 10.1128/JB.180.21.5540-5546.1998
[29] Kleerebezem M, Boekhorst J, Van Kranenburg R, et al. Complete genome sequence of Lactobacillus plantarum WCFS1[J]. Proceedings of the National Academy of Sciences of the United States of America,2003,100(4):1990−1995. doi: 10.1073/pnas.0337704100
[30] Padan E, Bibi E, Ito M, et al. Alkaline pH homeostasis in bacteria: New insights[J]. Biochimica Et Biophysica Acta Biomembranes,2005,1717(2):67−88. doi: 10.1016/j.bbamem.2005.09.010
[31] Wang Y, Chen C, Ai L, et al. Complete genome sequence of the probiotic Lactobacillus plantarum ST-III[J]. Journal of Bacteriology,2011,193(1):313−324. doi: 10.1128/JB.01159-10
[32] Chen C, Ai L, Zhou F, et al. Complete nucleotide sequence of plasmid pST-III from Lactobacillus plantarum ST-III[J]. Plasmid,2011,67(3):236−244.
[33] 乌日娜, 宋雪飞, 刘倩颖, 等. 植物乳杆菌分子伴侣蛋白基因在盐胁迫下的表达分析[J]. 食品科学,2015(11):119−123. doi: 10.7506/spkx1002-6630-201511023 [34] Cotter P D, Hill C. Surviving the acid test: Responses of gram-positive bacteria to low pH[J]. Microbiology and Molecular Biology Reviews,2003,67(3):429−442. doi: 10.1128/MMBR.67.3.429-453.2003
[35] Liu S W, Li K, Yang S L, et al. Development of a SCAR (Sequence-characterised amplified region) marker for acid resistance-related gene in Lactobacillus plantarum[J]. Extremophiles,2015,19(2):355−361. doi: 10.1007/s00792-014-0721-2
[36] Derzelle S, Hallet B, Francis K P, et al. Changes in cspL, cspP, and cspC mRNA abundance as a function of cold shock and growth phase in Lactobacillus plantarum[J]. Journal of Bacteriology,2000,182(18):5105−5113. doi: 10.1128/JB.182.18.5105-5113.2000
[37] Castaldo C, Siciliano R A, Muscariello L, et al. CcpA affects expression of the groESL and dnaK operons in Lactobacillus plantarum[J]. Microbial Cell Factories,2006,5(1):35−35. doi: 10.1186/1475-2859-5-35
[38] 刘倩颖. 基于RT-PCR技术对植物乳杆菌耐盐分子机理的研究[D]. 哈尔滨: 东北农业大学, 2014. [39] Eric M, Francoise B, Esther I, et al. Comparative proteomic analysis of Lactobacillus plantarum for the identification of key proteins in bile tolerance[J]. Bmc Microbiology,2011,11(1):63−63. doi: 10.1186/1471-2180-11-63
[40] Engelhardt T, Albano H, Kisko G, et al. Antilisterial activity of bacteriocinogenic Pediococcus acidilactici HA6111-2 and Lactobacillus plantarum ESB 202 grown under pH and osmotic stress conditions[J]. Food Microbiology,2015,48:109−115. doi: 10.1016/j.fm.2014.11.015
[41] Lim S M. Cultural conditions and nutritional components affecting the growth and bacteriocin production of Lactobacillus plantarum KC21[J]. Food Science & Biotechnology,2010,19(3):793−802.
[42] Leal-Sanchez M V, Jimenez-Diaz R, Maldonado-Barragan A, et al. Optimization of bacteriocin production by batch fermentation of Lactobacillus plantarum LPCO10[J]. Applied & Environmental Microbiology,2002,68(9):4465−4471.
[43] Vazquez J A, Cabo M L, Gonzalez M P, et al. The role of amino acids in nisin and pediocin production by two lactic acid bacteria: A factorial study[J]. Enzyme & Microbial Technology,2004,34(3):319−325.
[44] Yi H, Han X, Yang Y, et al. Effect of exogenous factors on bacteriocin production from Lactobacillus paracasei J23 by using a resting cell system[J]. International Journal of Molecular Sciences,2013,14(12):24355−24365. doi: 10.3390/ijms141224355
[45] Parlindungan E, Dekiwadia C, Tran K T M, et al. Morphological and ultrastructural changes in Lactobacillus plantarum B21 as an indicator of nutrient stress[J]. Lwt-Food Science and Technology,2018,92:556−563. doi: 10.1016/j.lwt.2018.02.072
[46] Cortes B W, Naditz A L, Anast J M, et al. Transcriptome sequencing of Listeria monocytogenes reveals major gene expression changes in response to lactic acid stress exposure but a less pronounced response to oxidative stress[J]. Frontiers in Microbiology,2020,10:3110. doi: 10.3389/fmicb.2019.03110
[47] Zheng S, Sonomoto K. Diversified transporters and pathways for bacteriocin secretion in gram-positive bacteria[J]. Applied Microbiology and Biotechnology,2018,102(10):4243−4253. doi: 10.1007/s00253-018-8917-5
[48] Ushijima Y, Ohniwa R L, Morikawa K. Identification of nucleoid associated proteins (NAPs) under oxidative stress in Staphylococcus aureus[J]. Bmc Microbiology,2017,17:8. doi: 10.1186/s12866-016-0922-1
[49] Froderberg L, Houben E N G, Baars L, et al. Targeting and translocation of two lipoproteins in Escherichia coli via the SRP/Sec/YidC pathway[J]. Journal of Biological Chemistry,2007,279(30):31026.
[50] Neumann-Haefelin C, Schafer U, Muller M, et al. SRP-dependent co-translational targeting and SecA-dependent translocation analyzed as individual steps in the export of a bacterial protein[J]. Embo Journal,2014,19(23):6419−6426.
[51] Lin J T, Zhu Y F, Tang H L, et al. Identification of a GntR family regulator BusR(Tha) and its regulatory mechanism in the glycine betaine ABC transport system of Tetragenococcus halophilus[J]. Extremophiles,2019,23(4):451−460. doi: 10.1007/s00792-019-01096-6
[52] Tsirigotaki A, De Geyter J, Sostaric N, et al. Protein export through the bacterial Sec pathway[J]. Nature Reviews Microbiology,2017,15(1):21−36. doi: 10.1038/nrmicro.2016.161
[53] 宋雪飞, 郭晶晶, 姜静, 等. 植物乳杆菌FS5-5在盐胁迫下的转录组学分析[J]. 食品科学,2017(6):26−32. [54] 张明阳. argG、argH和argR基因对Lactococcus lactis NZ9000胁迫抗性的影响[D]. 无锡: 江南大学, 2016. [55] Vrancken G, Rimaux T, Wouters D, et al. The arginine deiminase pathway of Lactobacillus fermentum IMDO 130101 responds to growth under stress conditions of both temperature and salt[J]. Food Microbiology,2009,26(7):720−7274. doi: 10.1016/j.fm.2009.07.006
[56] Heunis T, Deane S, Smit S, et al. Proteomic profiling of the acid stress response in Lactobacillus plantarum 423[J]. Journal of Proteome Research,2014,13(9):4028−4039. doi: 10.1021/pr500353x
[57] Jeon E, Lee S, Won J I, et al. Development of Escherichia coli MG1655 strains to produce long chain fatty acids by engineering fatty acid synthesis (FAS) metabolism[J]. Enzyme & Microbial Technology,2011,49(1):44−51.
[58] 王茜茜, 宋雪飞, 郭晶晶, 等. 基于iTRAQ技术对植物乳杆菌FS5-5的耐盐特性分析[J]. 微生物学报,2017,57(10):1461−1470. [59] Pieterse B. Unravelling the multiple effects of lactic acid stress on Lactobacillus plantarum by transcription profiling[J]. Microbiology,2005,151(12):3881−3894. doi: 10.1099/mic.0.28304-0
[60] Sun Y, Fukamachi T, Saito H, et al. ATP requirement for acidic resistance in Escherichia coli[J]. Journal of Bacteriology,2011,193(12):3072−3077. doi: 10.1128/JB.00091-11
[61] Pang B, Mcfaline J L, Burgis N E, et al. Defects in purine nucleotide metabolism lead to substantial incorporation of xanthine and hypoxanthine into DNA and RNA[J]. Proceedings of the National Academy of Sciences of the United States of America,2012,109(7):2319−24. doi: 10.1073/pnas.1118455109
[62] Hormann S, Scheyhing C, Behr J, et al. Comparative proteome approach to characterize the high-pressure stress response of Lactobacillus sanfranciscensis DSM 20451T[J]. Proteomics,2010,6(6):1878−1885.
[63] Vanbogelen R A, Neidhardt F C. Ribosomes as sensors of heat and cold shock in Escherichia coli[J]. Proceedings of the National Academy of Sciences of the United States of America,1990,87(15):5589−5593. doi: 10.1073/pnas.87.15.5589
[64] 陈卫, 赵山山, 张秋香. 乳酸菌的耐盐机制[J]. 中国食品学报,2013,13(10):1−7. [65] Kovacic L, Paulic N, Leonardi A, et al. Structural insight into LexA-RecA interaction[J]. Nucleic Acids Research,2013,41(21):9901−9910. doi: 10.1093/nar/gkt744
 下载:
下载:



 下载:
下载:

计量
- 文章访问数: 365
- HTML全文浏览量: 201
- PDF下载量: 19
