


Research Progress on the Effects of Processing Methods of Flavonoids in Tartary Buckwheat
-
摘要: 苦荞是传统的食药两用资源,含丰富的营养与功能成分,具有降糖、降脂、降压和抗炎等多种活性,已被开发成各类食品。黄酮类化合物是苦荞中主要的生物活性物质,对人体健康具有促进作用。芦丁和槲皮素是苦荞黄酮类化合物最主要的存在形式,不同加工方式对其含量及转化有显著影响,从而影响苦荞及其相关制品的功能活性和口感;然而目前缺乏相关的系统论述,不利于差异化苦荞健康产品的开发。本文综述了苦荞中黄酮类成分的稳定性和生物转化特性,以及制粉、萌发、发酵、热处理等几种常见加工方式对苦荞中黄酮类成分的影响规律,对苦荞加工过程中黄酮类成分的保持与转化具有重要指导作用,为开发针对性强的苦荞健康制品提供参考。Abstract: Tartary buckwheat is a traditional dual-use food and medicine resource. It is rich in nutrients and functional ingredients. It has various activities such as reducing blood sugar, reducing blood fat, reducing blood pressure and anti-inflammatory. It has been developed into various foods. Flavonoids are the main bioactive substances in Tartary buckwheat which promote human health. Among them, rutin and quercetin are the main existing forms of Tartary buckwheat flavonoids, different processing methods have a significant effect on its content and transformation, thus affecting the functional activity and taste of Tartary buckwheat and its related products. However, the lack of relevant systematic discussion is not conducive to the development of differentiated Tartary buckwheat health products.This artical reviews the stability and biotransformation characteristics of flavonoids in Tartary buckwheat, and the influence of several common processing methods on flavonoids, such as crushing, germination, fermentation and heat treatment. It plays an important role in the maintenance and transformation of flavonoids in Tartary buckwheat, and provides a reference for the development of health products with strong pertinence.
-
Keywords:
- Tartary buckwheat flavonoids /
- crushing /
- germination /
- fermentation /
- heat treatment
-
荞麦,又称三角麦,乌麦,灰皮麦,泛指来源于蓼科(Polygonaceae)荞麦属(Fagopyrum)植物荞麦F. esculentum Moench、苦荞麦F. tataricum(L.)Gaertn或翅荞麦F. emarginatum Mtissner等的果实[1],苦荞麦F. tataricum和荞麦F. esculentum是主要的栽培物种。我国苦荞麦资源丰富,种植面积及产量全球第一,主要分布在西南丘陵山区,如四川凉山、贵州毕节等地。目前荞麦已知品种编目共有2785份,其中苦荞879份,甜荞1896,野生10份[2]。苦荞生长期较短,2~3个月即可成熟,耐干旱,抗贫瘠,容易种植[3- 4]。苦荞营养丰富全面,其中蛋白质含量10%~15%,高于大米、玉米等常见粮食作物;脂肪含量2.1%~2.6%;淀粉含量为65%~75%,其中抗性淀粉占7.5%~8.5%;膳食纤维含量1.5%~2.1%;苦荞中钙、磷、铜、锌含量是稻米和小麦的2~3倍;苦荞中含有丰富的维生素,如维生素B1,维生素B2,维生素B6,维生素E,以及其他谷物没有的维生素P[5]。苦荞中不饱和脂肪酸含量丰富,尤其是亚油酸和油酸[6-7]。
黄酮类化合物是植物在生长过程中为抵御恶劣生态环境而形成的一大类次生代谢产物[8],广泛分布于植物界中具有2-苯基色原酮结构的化合物[9]。黄酮类化合物是苦荞中最重要的生物活性物质[10],苦荞籽粒中的黄酮类化合物主要有:芦丁、槲皮素、山奈酚、槲皮素-3-芸香糖苷、槲皮素-3-葡萄糖芸香糖苷、槲皮素-3-芸香糖双葡萄糖苷[9, 11]。据统计,在所有栽培苦荞中,芦丁的平均含量为19.96 mg/g,占总黄酮的75%以上,槲皮素平均含量为0.65 mg/g,山奈酚-3-O-芸香糖苷的平均含量为1.87 mg/g[12]。不同品种之间成分含量存在差异,但总体来说,黄酮类化合物的含量保持在较高水平。通过体内实验和流行病学调查,苦荞对人体健康具有很多积极的影响:苦荞中各成分协同作用,对人体DNA具有保护作用[13],同时具有抗癌、抗炎[14]、抗氧化、降血压[15]、抗疲劳[16]、降血糖[17]、改善阿尔兹海默症、保护神经元[18]、抗菌等活性,能降低胆固醇和低密度脂蛋白[19],对肝损伤具有保护作用[20]。另外,作为一类弱雌性激素,黄酮类化合物在妇女更年期综合征的治疗方面有显著作用[8]。苦荞黄酮类化合物对于改善大鼠的高脂血症有较好作用,能改善糖耐量,对于大肠杆菌、金黄色葡萄球菌、枯草芽孢杆菌、沙门氏菌具有明显的抑制作用[10, 21-22];大部分毒理学研究提示黄酮类化合物安全性高,作为食品和药品原材料是可行的[23]。
苦荞籽粒中含有高活性的蛋白酶抑制剂、芦丁降解酶、抗性淀粉等抗营养因子,这些物质的存在会降低苦荞籽粒的营养价值[24],粉碎、萌动、发酵、热处理等是苦荞加工过程中较为常见的处理方法,可以影响苦荞黄酮类化合物的种类及含量,改善苦荞及其制品的特性和口感。不同的加工方式对苦荞黄酮类化合物的影响具有很大的差异,本文分析了常规加工方法对苦荞及其制品中黄酮类化合物的影响,旨在更好的指导苦荞加工,为苦荞精准化营养产品开发提供参考。
1. 苦荞中的黄酮类化合物分布及其稳定性
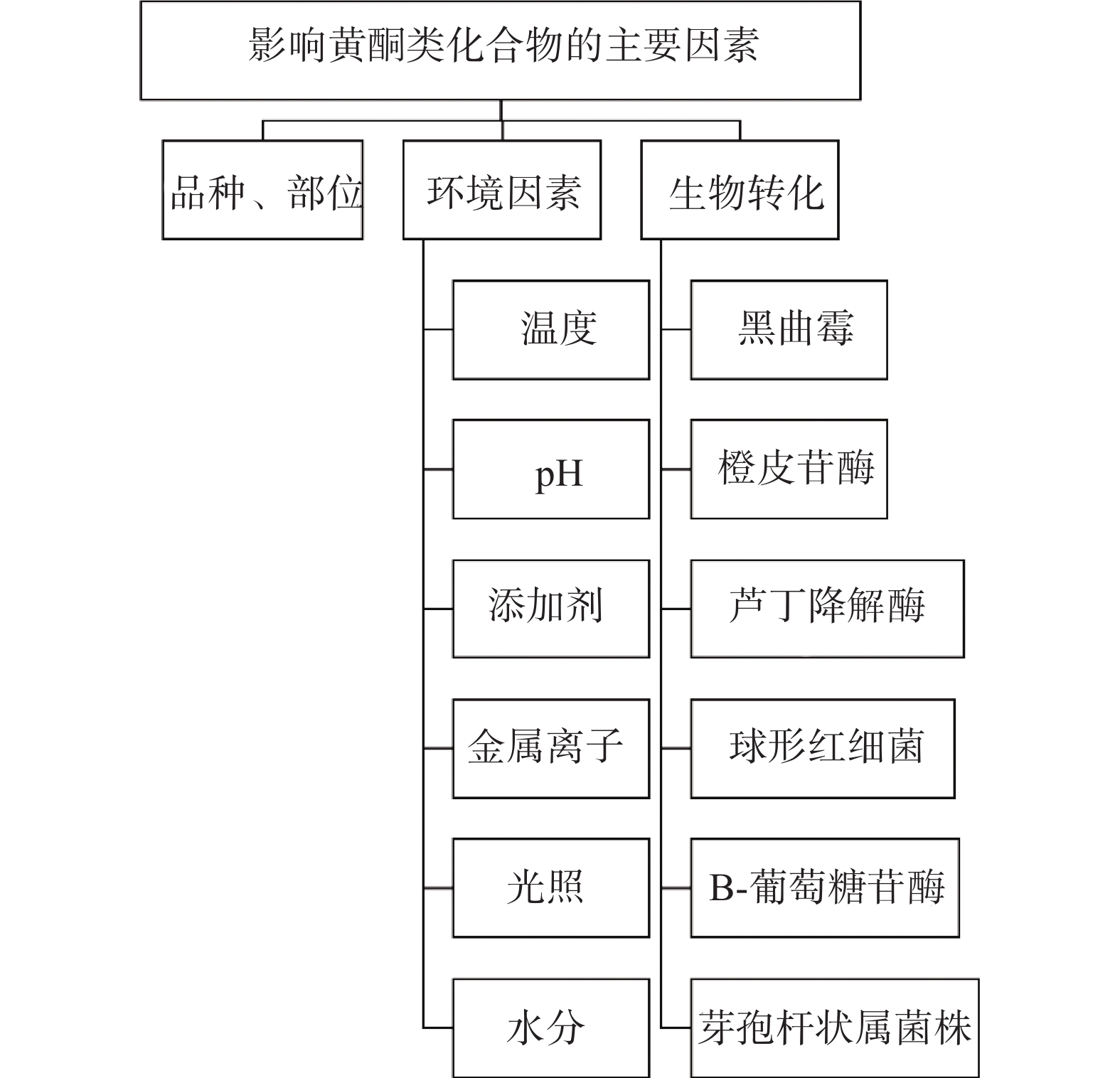
苦荞籽粒中富含黄酮类化合物,其总黄酮含量是甜荞麦的10~100倍。如图1所示,苦荞中不同品种、部位黄酮类化合物的含量差异、黄酮类化合物的稳定性及其生物转化特点是影响苦荞品质的重要因素,在加工时应重点考虑。
1.1 不同品种、部位黄酮类化合物含量差异
植物体内黄酮类化合物的含量和分布因品种、器官和生长期等的不同而存在差异。不同品种的黄酮类化合物组成与含量存在较大差异,同一品种在不同生长期、不同器官中的含量也有明显差异。课题组前期测定了53份不同生态区域、不同品种苦荞中芦丁、槲皮素、山奈酚的含量,结果表明苦荞籽粒中以芦丁含量最高,不同品种苦荞中芦丁含量最低为9.94 mg/g,最高为18.55 mg/g,说明不同品种苦荞品质差异较大。同一产地不同品种苦荞、同一品种苦荞在不同生态区种植,其总黄酮含量差异明显[25-26];苦荞芽在生长期,叶的总黄酮含量明显高于茎和根,苦荞籽粒外层粉的总黄酮含量最高,为5.23%~7.43%,芯粉最低,为0.47%~0.975%[9- 10]。在加工时选择不同品种或不同部位都会影响产品中黄酮类化合物的含量。
1.2 黄酮类化合物的稳定性
植物储存和加工过程中黄酮类化合物的稳定性受温度、添加剂、pH、金属离子等的影响。不同类型的植物中含有不同种类的黄酮类化合物,稳定性具有差异。研究显示,陈艾和蛹虫草中的总黄酮含量均随着储存时间的增加表现出先增加后降低的趋势[27-28],银杏叶总黄酮在光照条件下发生强烈的氧化和降解反应,结构破坏,总黄酮含量降低[29]。加热干燥后的三叶青块根中的总黄酮含量明显低于冷冻干燥和阴干,−20 ℃保存后总黄酮含量明显高于常温处理[30],采取不同的发汗方式对杜仲皮总黄酮影响不同,蒸煮后发汗与其他低温后发汗(如晒至半干后发汗,60 ℃烘至半干,阴至半干后发汗)相比,总黄酮含量明显降低[31]。太行菊在温度高于50 ℃的条件下储存,总黄酮稳定性变差[32],银杏叶总黄酮在温度高于80 ℃会被破坏[28],苦荞黄酮类化合物在温度高于90 ℃会逐渐分解[33],高温会使植物细胞的活性成分失活或活性降低,而低温有利于总黄酮的保留,在加工和储存过程中应该避免高温处理[29, 31]。煮制和蒸制过程中黄酮含量会因为是否与水直接接触而表现出差异,在煮制过程中总黄酮溶于水中而流失,并且在高温条件下发生水解、裂解等反应[33]。研究发现,添加剂对总黄酮也存在影响,苯甲酸钠会使蜂胶黄酮类化合物产生黄色絮状物影响黄酮类化合物活性,而对银杏叶总黄酮的影响不明显[29, 34],在加工时应该根据不同黄酮类化合物的性质而适宜添加。黄酮类化合物因含有多个酚羟基呈现弱酸性,在强碱性环境下表现出较差的稳定性,在pH=5~7的条件下稳定性较好[29, 31-32]。金属离子Ca2+、Al3+、Fe2+、Fe3+等会与黄酮类化合物结构中的邻二酚羟基发生络合反应,明显影响黄酮类化合物的稳定性,还原剂NaSO3也会降低黄酮类化合物的含量。综上,在植物储藏过程中,应避免使用含有SO2、亚硫酸盐、及Ca2+、Al3+、Fe2+、Fe3+等金属离子的容器和材料,避免强酸强碱环境,在阴凉、避光处保存,最大限度保留黄酮类化合物含量[32, 35]。
1.3 黄酮类化合物的生物转化
近年来,对黄酮类化合物生物转化的研究越来越多。在植物的生长加工过程中,影响其含量的因素很多。天然黄酮类化合物主要以苷类形式存在,由于苷元、连接苷元的糖的种类、数量、位置等不同而形成了不同的黄酮苷类化合物,通过生物转化技术可以增加黄酮类化合物的含量和活性。黑曲霉作为世界公认的食用安全性的低等真核生物,在黑曲霉产生的丰富酶系的参与下进行酶促反应,可以促进黄酮类化合物的转化[36]。陈皮受黑曲霉的污染,由于受其代谢活动的调节,陈皮中的黄酮类化合物含量增加[37]。柚皮苷在黑曲霉参与的条件下产生柚苷酶液使柚皮苷发生去糖基化生成柚皮素,可以改善柚皮苷的苦味[38]。芦丁在黑曲霉产生的鼠李糖酶的催化下发生去糖基化反应,转换为槲皮素-3-葡萄糖苷。橙皮苷在橙皮苷酶的处理下,转化成橙皮素-7-葡萄糖苷,橙皮素和柚皮素的抗氧化活性均强于橙皮苷和柚皮苷[39]。在浸泡和加热的条件下,β-葡萄糖苷酶活性的增加对于飞燕草素和花青素黄酮苷元的积累有促进作用[40]。金莲花的黄酮苷类化合物在肝微粒体中发生羟基化、乙酰化、糖基化等代谢反应,使黄酮类化合物相互转化[41],增强了黄酮类化合物的药效,更易吸收。槲寄生提取物经过球形红细菌转化后,总黄酮含量增加129%[42],藜蒿的总黄酮含量经芽孢杆菌属菌株生物转化后增加了7.56倍[43]。
研究显示,苦荞粉与水接触30 s时已有55.2%的芦丁被分解,3 min后下降趋于平缓,而槲皮素的含量与芦丁刚好相反,反映了芦丁向槲皮素转换的过程。苦荞蒸煮处理40~60 min后,芦丁含量提高,推测在蒸煮一定时间后发生了槲皮素向芦丁转换的逆反应,结合Barber提出的芦丁合成途径:槲皮素先发生3-糖基化反应生成异槲皮素,异槲皮素再经鼠李糖基化反应生成芦丁,推测芦丁被芦丁降解酶水解为槲皮素的同时会发生芦丁的合成反应,但该推论尚未得到证实,需进行进一步的研究[44-45]。水的加入会使芦丁转换为槲皮素,槲皮素口感苦涩,在苦荞面食类产品制作时,应充分考虑芦丁保留的问题,选择适当的灭酶加工方法,减少芦丁向槲皮素的转化。如Bae等[46]用富含芦丁的苦荞粉(REFE)制作的面条可以降低苦味,其主要特点在于苦荞种子加工前进行了蒸煮处理,芦丁降解酶的失活可以解释REFE中槲皮素的缺失,这与Yoo[47]的报道一致。研究显示,添加富含芦丁或槲皮素的苦荞粉制作的面条,其淀粉消化率均显著下降,其中添加富含槲皮素苦荞粉的面条下降幅度更大,说明通过适当的工艺控制可生产不同活性的苦荞产品[48]。另外,造粒型苦荞茶和全籽粒型苦荞茶中的总黄酮含量存在显著差异,造粒型苦荞茶中富含槲皮素,而全籽粒型苦荞茶中富含芦丁,就是由于两者加工工艺不同造成。在针对不同人群食用时应注意槲皮素与芦丁活性的差异,利用苦荞中芦丁降解酶的存在控制芦丁和槲皮素的适当转化,提高营养“针对性”。综上,通过生物转化可以提高黄酮类化合物等次生代谢产物的种类和含量,并增加其稳定性、水溶性和生物利用度等,增强苦荞制品品质[49]。
2. 不同处理过程中苦荞黄酮类化合物的变化
2.1 制粉对苦荞黄酮类化合物的影响
相比清洗和较低温度的干燥处理,粉碎对苦荞黄酮类化合物有更大的影响。如表 1 所示,苦荞籽粒不同部位中黄酮类化合物含量差异显著,不同粒径的苦荞粉总黄酮含量差异较大。苦荞麸皮中富含功能物质和多种营养素,其总黄酮含量约为苦荞胚乳部位的4.86倍[50]。苦荞制粉过程一般将过120目的苦荞粉称为“芯粉”,而80~120目的用作“皮粉”。芯粉主要组成为苦荞胚乳部分,而皮粉中麸皮比例增加,因此其黄酮类化合物含量增加,口感更苦。随着粉碎程度的增加,总黄酮含量呈现先减小后增大趋势,粉碎过程中受不同程度的综合作用力,破坏了黄酮类化合物的结构;粉碎程度越高,平均粒径越小,粉体均匀性增加,使苦荞粉中总黄酮的提取更加充分,总黄酮含量略微增加[51]。超微粉碎处理能提高苦荞麸皮中的功能成分溶出,通过对比发现不同超微粉碎方式对苦荞粉在人工胃肠液中总黄酮溶出率存在的差异,气流超微粉碎的溶出得率极显著高于振动超微粉碎和万能粉碎[52],原因是粒径大小存在差异,同时气流粉碎中苦荞较大颗粒已经开裂促使黄酮类化合物较易溶出。
表 1 制粉过程对苦荞黄酮类化合物的影响Table 1. Effect of the crushing process on tartary buckwheat flavonoids加工方法 黄酮类化合物含量变化 参考文献 粉碎 游离黄酮类化合物:皮粉(918.47 mg/100 g)>
芯粉(272.7 mg/100 g);结合黄酮类化合物:
皮粉(252.11 mg/100 g)>芯粉(121.71 mg/100 g)[50] 随粉碎程度的增加,4种不同粒径的苦荞粉(d(0.5)
分别为33.051、19.763、16.538、15.289 μm)总黄酮含
量较普通粉(d(0.5)=103.601 μm)相比依次下降
7.92%、9.40%、17.59%、上升2.59%[51] 在人工胃肠液中苦荞总黄酮的溶出得率:
气流超微粉碎>振动超微粉碎>普通万能机粉碎,
在人工肠液中,第5 h时气流超微粉碎的溶出率是
普通万能机粉碎的2倍左右[52] 苦荞制粉过程是苦荞加工的首要步骤,加工参数的差异对苦荞营养物质的影响不同。苦荞中富含芦丁降解酶,芦丁主要存在于胚乳,而芦丁降解酶主要存在于种皮中,两者的空间位阻在一定程度上限制了芦丁降解酶的作用,苦荞粉碎后,两者在少量水存在的状态下迅速反应,造成芦丁含量的下降[26]。苦荞制粉前如果增加灭酶环节(如蒸汽灭酶),则可在后续加工过程中避免芦丁向槲皮素的转化,反之,芦丁在苦荞粉接触水后迅速转化成槲皮素。利用该性质可以控制芦丁和槲皮素的转化,最大限度保留芦丁,从而改善苦荞制品口感,或生产富含芦丁/槲皮素的功能差异化产品。
2.2 萌发过程中苦荞黄酮类化合物的变化
苦荞萌发过程可以提高并改善苦荞籽粒的活力,提高相关酶活力及生物活性成分的含量,在此过程中,苦荞黄酮类化合物的含量受多种因素的影响,如:苦荞黄酮类化合物代谢相关酶、浸种时间、温度、光照、微量元素等[9]。在整个萌发过程中,苦荞籽粒中黄酮类化合物的含量均有明显增加,原因是在浸泡萌发的过程中,大量结合态的黄酮类化合物被酶水解而释放出来[53]。黄酮类化合物的合成是受多种关键酶共同作用影响,在特定酶的作用下,植物会向着某一特定黄酮类化合物的方向进行合成。苯丙氨酸解氨酶(PAL)是黄酮类化合物合成过程中的第一个关键性酶[54-55],查尔酮异构酶(CHI)是黄酮类次级代谢产物积累的关键酶,降低或抑制PAL或CHI的表达活性都会影响黄酮类化合物的积累。对于种子的处理条件不同,PAL酶活性峰值出现的时间也有明显差异,当酶活力达到峰值时,黄酮类化合物的含量增长最快。在萌芽前期,黄酮类化合物含量的增加受PAL和CHI共同作用导致,但是后期主要受CHI活性的影响。研究显示,PAL和CHI在发芽的苦荞中的活性与总黄酮含量呈现正相关[56]。
如表 2 所示,在苦荞萌芽过中,黄酮类化合物含量受环境条件(温度、光照、磁场)和外源添加物(微量元素、NaCl、苯丙氨酸、香菇多糖)等的影响[9, 57-58]。在不同的萌发条件下,黄酮类化合物变化存在差异。苦荞喜阴凉的生长环境,在25 ℃条件下萌发有最高的总黄酮含量[56],22 ℃条件下萌发芦丁含量最高[59],不同物质代谢途径不同,导致最佳萌发温度存在一定的差异。苦荞组织对不同光质的敏感度不同,调控总黄酮积累的基因,如Ft4CL(4-香豆酞-CoA连接酶)的表达量在紫外条件下显著上升,因此紫外照射组相比黑暗组总黄酮含量上升显著[60]。不同光周期处理,调控总黄酮合成相关基因的表达量存在差异,在16 h/d的光周期处理下,黄酮醇合成支路关键酶基因FtFLS1表达量上升,且显示出强烈的光诱导特征,芦丁含量第四天有最高水平,并且充足的光照时间使植物积累更多的碳水化合物,有利于其生长发育[61]。不同的磁场强度条件下,不同酶的诱导效应不同,当磁场强度为0.3 T时,PAL、CHI、RDEs3种酶协同作用诱导总黄酮的积累,含量达到最高值 (62.90 mg/g) [58]。外源添加物也会导致黄酮类化合物含量的变化,已有研究表明Al3+对大多数黄酮类化合物具有抑制作用,但在苦荞萌发过程中施加不同的Al3+离子溶液,能够显著增加总黄酮的含量,这与Al3+改善苯丙氨酸代谢关键酶:PAL和CHI的活性密切相关。金属离子作为代谢的关键媒介,会影响代谢途径或刺激植物的自卫机制[56]。苦荞对铝有解毒机制,槲皮素是金属解毒剂,在Al3+加入后,苦荞通过螯合铝从根部分泌黄酮类化合物和草酸来应对铝胁迫,刺激种子合成黄酮类化合物。超声协同NaCl胁迫处理能够提高黄酮类化合物合成相关酶基因的表达量,但NaCl浓度到达一定范围后,Na+与种子萌发所需要的酶发生反应或与酶特异结合,产生刺激物质或使酶活性降低而抑制苦荞的萌发,降低发芽率[62]。外源苯丙氨酸和香菇多糖的添加可以促进苯丙烷类代谢途径中PAL和CHI基因的表达,从而提高总黄酮含量[63-64]。综上可知,黄酮类化合物的积累受到多种因素的影响,萌发过程主要通过促进黄酮类化合物代谢相关酶的表达,从而促进黄酮类化合物的积累,萌发过程有利于丰富苦荞籽粒中营养物质的种类和数量。
表 2 萌发过程对苦荞黄酮类化合物的影响Table 2. Effect of germination process on tartary buckwheat flavonoids处理过程 黄酮类化合物含量变化 参考文献 萌发过程 总黄酮和芦丁含量均在萌发的第 11 d逐渐升高至最大值,之后处于一个相对稳定的水平,其中,第 11 d时总黄酮为51.50 mg/g,芦丁含量为 47.96 mg/g,总黄酮较未经萌发的苦荞种子增加了2.4倍。 [56] 不同磁场强度条件下萌发 当磁场强度为3 T时,总黄酮含量达到最高值62.90 mg/g [58] 不同温度条件下萌发 25 ℃条件下萌发有最高的总黄酮含量(1090.25 mg/100 g),22 ℃条件下萌发芦丁含量最高(105 mg/100 g) [56, 59] 不同光质条件下萌发 子叶的总黄酮含量在紫外光(385 nm)照射条件下最高,相比对照组增加了2.08倍;LED蓝光(460 nm)次之,为对照组的1.24倍;红光(630 nm)和白光(540 nm)对子叶总黄酮的影响相对较小 [60] 不同光周期条件萌发 16 h/d的光周期处理下,芦丁含量在第4 d时达到最高(59.60 mg/g),显著高于整个萌芽过程中的平均芦丁含量(46.47 mg/g),之后逐渐下降 [61] 施加不同的Al3+离子溶液 随着Al3+浓度的增加,总黄酮含量上升,当Al3+浓度为1000 mg/L 时,总黄酮含量达到最大值(1315.52 mg/100 g),继续增加浓度,总黄酮含量略有下降 [56] 超声波及NaCl胁迫处理 超声协同NaCl胁迫处理后,总黄酮含量明显高于对照;超声波处理21 min、超声波功率320 W、3.0 mmol/L的NaCl溶液处理条件下,苦荞的总黄酮含量明显增加,高达1.59 g/100 g,然而NaCl浓度过高会降低发芽率 [62] 外源苯丙氨酸的添加 添加外源苯丙氨酸后,第7 d总黄酮的积累量最高达4.84 g/100 g,相比对照组增加了28.38% [63] 香菇多糖的添加 香菇多糖的浓度处于100~200 μg/mL范围内时总黄酮含量增加最明显, 100 μg/mL香菇多糖处理后总黄酮含量最高,较对照组增加了18.69%;当香菇多糖浓度大于400 μg/mL后,总黄酮的合成受到抑制 [64] 2.3 发酵过程中苦荞黄酮类化合物的变化
苦荞由于其丰富的营养物质被广泛运用于食品加工中,但其适口性差,通过微生物发酵可以改善其口感,并且可将苦荞中的大分子物质转换成人体更易于吸收的小分子物质,如氨基酸类、醇类、有机酸类等生物活性成分。目前对于苦荞的发酵制品研究也越来越多,不同的发酵状态对苦荞中黄酮类化合物的影响也有一定的差异。
如表 3 所示,在苦荞酒的酿造中,糖化是芦丁、槲皮素大量损失的阶段,该研究中,麦汁的发酵温度为45 ℃,糖化用水pH为5.2,而芦丁降解酶的最适pH在5.0左右,最适作用温度为50 ℃,这为芦丁降解酶的作用提供了最适的条件,并且该过程中有水的参与,造成了芦丁大量损失。添加酒花煮沸后,酒花中含有的芦丁释放,芦丁保留率略微升高[65-66]。在苦荞醪糟的加工工艺中,随着发酵时间的延长,大分子物质进一步分解成小分子物质,芦丁和槲皮素含量升高[67]。苦荞醋的酿造过程中,酒精发酵时,发酵液中酒精浓度的增加导致芦丁在发酵液中的溶解度增加;醋酸发酵阶段酒精浓度降低,芦丁的溶解度降低而析出进入残渣中,导致发酵液中芦丁含量下降,而残渣中上升[68]。总的来说,苦荞长时间的发酵状态不利于芦丁和槲皮素的存在,但研究显示,苦荞发酵制品具有很强的抗氧化能力,发酵过程中产生更多的小分子物质如:氨基酸类、醇类、有机酸类等生物活性成分,有益于人体健康,并且更易被人体吸收。在苦荞的发酵过程中,应该合理控制发酵条件,尽可能保留苦荞中的黄酮类化合物,开发出更多益于人体健康的苦荞发酵制品。
表 3 发酵过程对苦荞黄酮类化合物的影响Table 3. Effect of fermentation process on flavonoids of tartary buckwheat2.4 热处理过程中苦荞黄酮类化合物的变化
热加工是食品加工过程中最基本,最常用的加工方式,受热处理的影响,产品的营养成分会发生变化。如表 4 所示,孙丹等[53]的研究发现,高温和高压的状态均会导致苦荞中总黄酮含量降低,煮制对芦丁、槲皮素的影响最小[69],而油炸和烙制对其影响较大。蒸煮过程中温度在一定范围内升高会使芦丁降解酶的活性升高,加快芦丁向槲皮素的转化,温度升高到达一定范围时,芦丁降解酶活性下降,芦丁向槲皮素的转化速度减慢[70]。蒸制过程中内部结构破裂,蛋白质变性,淀粉糊化,与大分子物质结合的结合黄酮类化合物释放,结合黄酮类化合物含量短暂上升;继续加热时,结合黄酮类化合物受热分解含量降低;游离的黄酮类化合物随着水分渗出而呈现下降趋势[70]。煮制过程中,游离黄酮类化合物和芦丁的变化基本同蒸制,但结合黄酮类化合物持续下降,因为煮制过程受热面积大,直接与水接触,加热程度更大,结合黄酮类化合物一旦释放迅速被分解。焙烤时,总黄酮和芦丁均下降,这也导致了苦荞面包在烤制过程中颜色变暗黄绿色,芦丁为黄绿色,而槲皮素为深绿色物质,芦丁转换成槲皮素,因此颜色变暗[70]。挤压膨化是一个高温高压的过程,在该过程中,受温度的影响,温度越高,总黄酮和总酚物质损失越大,色泽越深,对苦荞制品的蒸煮特性影响越大[26, 71]。总之,高温状态不利于苦荞黄酮类化合物的保留,加热过程中温度越高,总黄酮损失越大,尽管高温抑制了芦丁降解酶的活性,但芦丁仍然有一定程度的下降,若期望最大限度保留苦荞中的黄酮类化合物,应合理控制热处理条件,避免高温高压带来的损失。
表 4 热处理过程对苦荞黄酮类化合物的影响Table 4. Effect of heat treatment on flavonoids of tartary buckwheat3. 结语及展望
苦荞制品加工工艺复杂,影响其品质因素众多。在苦荞的加工过程中,常见的加工方式有粉碎、萌发、发酵、热处理等,这些加工方式对苦荞营养成分的影响不一,不同加工参数均会对苦荞黄酮类化合物产生影响。粉碎过程作为加工的首要步骤对苦荞黄酮类化合物的影响较大,尤其是粒径的大小对苦荞黄酮类化合物的影响最为显著;萌发过程主要通过促进黄酮类化合物代谢相关酶(PAL/CHI)等的表达使苦荞黄酮类化合物的含量获得较大的提升,适当改变萌发条件,如温度、光照、光周期、金属离子浓度以及外源添加物浓度等可以不同程度的促进苦荞黄酮类化合物的积累;发酵过程中发酵条件如pH、温度等通常情况下接近于芦丁降解酶的最适作用范围,芦丁降解酶活性增加,使得芦丁被大量破坏从而含量降低,尽管发酵过程能够产生氨基酸类、醇类、有机酸类等益于人体健康的生物活性成分,但对于芦丁的破坏也不能被忽视;热加工处理过程中高温状态造成黄酮类化合物发生水解、裂解等反应,总黄酮含量显著降低,另一方面,制粉前进行高温处理(如:蒸汽灭酶)可以降低芦丁降解酶的活性,减少芦丁的损失。总之,加工工艺影响苦荞中黄酮类化合物的种类与数量,从而影响其品质与口感,未来苦荞产品开发中,应充分掌握苦荞中黄酮类化合物的变化规律,指导苦荞“精准化”产品的开发,促进苦荞产业健康发展。
-
表 1 制粉过程对苦荞黄酮类化合物的影响
Table 1 Effect of the crushing process on tartary buckwheat flavonoids
加工方法 黄酮类化合物含量变化 参考文献 粉碎 游离黄酮类化合物:皮粉(918.47 mg/100 g)>
芯粉(272.7 mg/100 g);结合黄酮类化合物:
皮粉(252.11 mg/100 g)>芯粉(121.71 mg/100 g)[50] 随粉碎程度的增加,4种不同粒径的苦荞粉(d(0.5)
分别为33.051、19.763、16.538、15.289 μm)总黄酮含
量较普通粉(d(0.5)=103.601 μm)相比依次下降
7.92%、9.40%、17.59%、上升2.59%[51] 在人工胃肠液中苦荞总黄酮的溶出得率:
气流超微粉碎>振动超微粉碎>普通万能机粉碎,
在人工肠液中,第5 h时气流超微粉碎的溶出率是
普通万能机粉碎的2倍左右[52]  下载: 导出CSV
下载: 导出CSV
表 2 萌发过程对苦荞黄酮类化合物的影响
Table 2 Effect of germination process on tartary buckwheat flavonoids
处理过程 黄酮类化合物含量变化 参考文献 萌发过程 总黄酮和芦丁含量均在萌发的第 11 d逐渐升高至最大值,之后处于一个相对稳定的水平,其中,第 11 d时总黄酮为51.50 mg/g,芦丁含量为 47.96 mg/g,总黄酮较未经萌发的苦荞种子增加了2.4倍。 [56] 不同磁场强度条件下萌发 当磁场强度为3 T时,总黄酮含量达到最高值62.90 mg/g [58] 不同温度条件下萌发 25 ℃条件下萌发有最高的总黄酮含量(1090.25 mg/100 g),22 ℃条件下萌发芦丁含量最高(105 mg/100 g) [56, 59] 不同光质条件下萌发 子叶的总黄酮含量在紫外光(385 nm)照射条件下最高,相比对照组增加了2.08倍;LED蓝光(460 nm)次之,为对照组的1.24倍;红光(630 nm)和白光(540 nm)对子叶总黄酮的影响相对较小 [60] 不同光周期条件萌发 16 h/d的光周期处理下,芦丁含量在第4 d时达到最高(59.60 mg/g),显著高于整个萌芽过程中的平均芦丁含量(46.47 mg/g),之后逐渐下降 [61] 施加不同的Al3+离子溶液 随着Al3+浓度的增加,总黄酮含量上升,当Al3+浓度为1000 mg/L 时,总黄酮含量达到最大值(1315.52 mg/100 g),继续增加浓度,总黄酮含量略有下降 [56] 超声波及NaCl胁迫处理 超声协同NaCl胁迫处理后,总黄酮含量明显高于对照;超声波处理21 min、超声波功率320 W、3.0 mmol/L的NaCl溶液处理条件下,苦荞的总黄酮含量明显增加,高达1.59 g/100 g,然而NaCl浓度过高会降低发芽率 [62] 外源苯丙氨酸的添加 添加外源苯丙氨酸后,第7 d总黄酮的积累量最高达4.84 g/100 g,相比对照组增加了28.38% [63] 香菇多糖的添加 香菇多糖的浓度处于100~200 μg/mL范围内时总黄酮含量增加最明显, 100 μg/mL香菇多糖处理后总黄酮含量最高,较对照组增加了18.69%;当香菇多糖浓度大于400 μg/mL后,总黄酮的合成受到抑制 [64]
下载: 导出CSV
表 3 发酵过程对苦荞黄酮类化合物的影响
Table 3 Effect of fermentation process on flavonoids of tartary buckwheat
下载: 导出CSV
-
[1] 任长忠, 赵钢. 中国荞麦学[M]. 北京: 中国农业出版社, 2015: 3−5. [2] 周冉冉, 陈茂彬, 张玉. 苦荞营养、功能和香气成分的研究进展[J]. 中国酿造,2018,37(12):12−15. doi: 10.11882/j.issn.0254-5071.2018.12.003 [3] 赵钢, 邹亮, 彭镰心, 等. 铅胁迫对苦荞生理特性的影响[J]. 江苏农业科学,2012,40(7):98−100. doi: 10.3969/j.issn.1002-1302.2012.07.035 [4] 王艳. 不同季节多年生苦荞的叶及其发酵茶品质变化研究[D]. 贵阳: 贵州师范大学, 2019. [5] 张余, 黄小敏, 刘昌敏, 等. 苦荞营养保健成分及其食品开发研究进展与展望[J]. 粮食与油脂,2019,32(8):12−14. doi: 10.3969/j.issn.1008-9578.2019.08.004 [6] Yang J, Gu Z, Zhu L, et al. Buckwheat digestibility affected by the chemical and structural features of its main components[J]. Food Hydrocolloids,2019,96(11):596−603.
[7] 何伟俊, 曾荣, 白永亮, 等. 苦荞麦的营养价值及开发利用研究进展[J]. 农产品加工,2019(23):69−75. [8] 崔强, 陈景超. 黄酮类化合物生理活性及合成研究进展[J]. 黑龙江科技信息,2011(20):30−30. [9] 杨延利. 萌发对苦荞黄酮合成的影响及萌发物抑菌、抗肿瘤活性的研究[D]. 上海: 上海师范大学, 2011. [10] 秦培友. 我国主要荞麦品种资源品质评价及加工处理对荞麦成分和活性的影响[D]. 北京: 中国农业科学院, 2012. [11] 孙琳, 袁杰彬, 陈双为, 等. 苦荞中黄酮类化合物提取方法的研究进展[J]. 酿酒科技,2018(9):74−80. [12] 吕惠生, 冯永鑫, 周锦怡, 等. 苦荞活性组分超临界流体工业色谱提纯工艺[J/OL]. 高效化学工程学报, 2019, http://kns.cnki.net/kcms/detail/33.1141.TQ.20191024.0929.002.html. [13] Vogrincic M, Kreft I, Filipic M, et al. Antigenotoxic effect of tartary (Fagopyrum tataricum) and common (Fagopyrum esculentum) buckwheat flour[J]. J Med Food,2013,16(10):944−952. doi: 10.1089/jmf.2012.0266
[14] Karki R, Park C H, Kim D W. Extract of buckwheat sprouts scavenges oxidation and inhibits proinflammatory mediators in lipopolysaccharide-stimulated macropHages (RAW264.7)[J]. J Integr Med,2013,11(4):246−252. doi: 10.3736/jintegrmed2013036
[15] Tsai H, Deng H, Tsai S, et al. Bioactivity comparison of extracts from various parts of common and tartary buckwheats: Evaluation of the antioxidant- and angiotensin-converting enzyme inhibitory activities[J]. Chemistry Central Journal,2012,6(1).
[16] Jin H M, Wei P. Anti-fatigue properties of tartary buckwheat extracts in mice[J]. Int J Mol Sci,2011,12(8):4770−4780. doi: 10.3390/ijms12084770
[17] Qin P, Wu L, Yao Y, et al. Changes in pHytochemical compositions, antioxidant and α-glucosidase inhibitory activities during the processing of tartary buckwheat tea[J]. Food Research International,2013,50(2):562−567. doi: 10.1016/j.foodres.2011.03.028
[18] Choi J Y, Cho E J, Lee H S, et al. Tartary buckwheat improves cognition and memory function in an in vivo amyloid-beta-induced alzheimer model[J]. Food Chem Toxicol,2013,53:105−111. doi: 10.1016/j.fct.2012.11.002
[19] Yang N, Li Y M, Zhang K, et al. Hypocholesterolemic activity of buckwheat flour is mediated by increasing sterol excretion and down-regulation of intestinAl NPC1L1 and ACAT2[J]. Journal of Functional Foods,2014,6:311−318. doi: 10.1016/j.jff.2013.10.020
[20] Lee C C, Shen S R, Lai Y J, et al. Rutin and quercetin, bioactive compounds from tartary buckwheat, prevent liver inflammatory injury[J]. Food Funct,2013,4(5):794−802. doi: 10.1039/c3fo30389f
[21] 李栋, 张立攀, 李向力, 等. 苦荞降血糖产品的稳定性研究[J]. 河南科学,2020,38(1):57−62. doi: 10.3969/j.issn.1004-3918.2020.01.010 [22] Kai C, Qiang W, Shiqin W, et al. The facilitating effect of tartary buckwheat flavonoids and lactobacillus plantarum on the growth performance, nutrient digestibility, antioxidant capacity, and fecal microbiota of weaned piglets[J]. Animals: An Open Access Journal from MDPI, 2019, 9(11): 986.
[23] 龙海荣, 杨洋, 刘绍州, 等. 黄酮类化合物的安全性研究进展[J]. 食品研究与开发,2008(10):154−157. doi: 10.3969/j.issn.1005-6521.2008.10.047 [24] 张冬梅. 苦荞芽菜培养及其次生代谢产物黄酮的代谢调控[D]. 西安: 陕西科技大学, 2012. [25] 彭镰心. 基于调节糖脂代谢效应的苦荞资源品质研究[D]. 成都: 成都中医药大学, 2019. [26] 孙晓静. 糊化处理对苦荞面团性质的影响[D]. 咸阳: 西北农林科技大学, 2016. [27] 龚敏, 卢金清. 不同年份陈艾的总黄酮、总多糖研究[J]. 湖北农业科学,2019,58(13):114−116,121. [28] 李建平, 张铁, 曾文波. 不同储存时间对蛹虫草中化学成分含量的影响[J]. 北方园艺,2019(22):134−142. [29] 冯靖, 彭效明, 李翠清, 等. 银杏叶黄酮的抗氧化性及其稳定性研究[J]. 食品科技,2019,44(4):244−249. [30] 洪春桃, 沈登锋, 魏斌, 等. 干燥与储存方式对三叶青块根总黄酮含量的影响[J]. 浙江农业科学,2019,60(12):2316−2318. [31] 李雪营, 林先燕, 孙晓惠, 等. 杜仲不同“发汗”加工方法制品中总黄酮含量的比较[J]. 时珍国医国药,2019,30(3):597−599. [32] 葛水莲, 陈建中, 刘娜, 等. 太行菊总黄酮抗氧化活性及稳定性研究[J]. 食品科技,2019,44(10):241−245. [33] 王丽娟, 刘苏萌, 李可, 等. 黑苦荞中黄酮类化合物的稳定性和抑菌活性研究[J]. 粮食与油脂,2015,28(8):62−65. doi: 10.3969/j.issn.1008-9578.2015.08.017 [34] 黄健, 郑敏, 张志鹏. 5种食品添加剂对蜂胶总黄酮稳定性的影响[J]. 湖北科技学院学报(医学版),2019,33(5):374−376,461. [35] 汪道兵, 高青海, 孙玉军. 老鸦瓣黄酮的制备及其稳定性研究[J]. 江汉大学学报(自然科学版),2019,47(6):549−554. [36] 王智磊, 刘素娟, 张鑫, 等. 黑曲霉生物转化黄酮类成分研究进展[J]. 中国实验方剂学杂志,2017,23(21):220−228. [37] 王福, 张鑫, 卢俊宇, 等. 陈皮“陈久者良”之黄酮类成分增加原因探究[J]. 中国中药杂志,2015,40(24):4890−4896. [38] 邓媛, 毛勇, 王燕. 黑曲霉 TC-01 产柚苷酶对柚皮苷酶解作用的研究[J]. 中国食品添加剂,2012(3):108−111. doi: 10.3969/j.issn.1006-2513.2012.03.015 [39] Da S C M G, Contesini F J, Sawaya A, et al. Enhancement of the antioxidant activity of orange and lime juices by flavonoid enzymatic de-glycosylation[J]. Food Research International,2013,52(1):308−314. doi: 10.1016/j.foodres.2013.03.019
[40] F. H Y H H J. The conversion and deglycosylation of isoflavones and anthocyanins in black soymilk process[J]. Food Chemistry,2018,261:8−14. doi: 10.1016/j.foodchem.2018.03.152
[41] 刘双月, 安燕南, 刘斯琪, 等. 金莲花中8种黄酮类成分的肝微粒体生物转化研究[J]. 中国现代中药,2019,21(11):1489−1496. [42] 郑庆红, 张忠鹏, 耿子颖, 等. 球形红细菌生物转化槲寄生中总黄酮类化合物的测定[J]. 中国药物与临床,2012,12(8):981−983. doi: 10.3969/j.issn.1671-2560.2012.08.001 [43] 李宁, 叶子茂, 向福, 等. 藜蒿总黄酮生物转化菌株的筛选及转化工艺优化[J]. 中国酿造,2018,37(6):150−154. [44] Barber G, Behrman E J. The synthesis and characterization of uridine 50-(b-L-rhamnopyranosyl dipHospHate) and its role in the enzymatic synthesis of rutin[J]. Archives of Biochemistry and BiopHysics,1991,288:239−242.
[45] Li D, Li X L, Ding X L, et al. A process for preventing enzymatic degradation of rutin in tartary buckwheat (Fagopyrum tataricum Gaertn) flour[J]. Food Science and Biotechnology,2008,17:118−122.
[46] Bae I Y, Choi A S, Lee H G. Impact of buckwheat flavonoids on in vitro starch digestibility and noodle-making properties[J]. Cereal Chemistry Journal,2016,93(3):299−305. doi: 10.1094/CCHEM-03-15-0047-R
[47] Yoo J Y, Kim, Y J, et al. Reduction of rutin loss in buckwheat noodles and their physicochemical characterisation[J]. Food Chem,2012,132:2107−2111. doi: 10.1016/j.foodchem.2011.12.065
[48] Zhang J, Wang F, Han P, et al. Effect of tartary buckwheat peptides on shelf life of tilapia (Oreochromis niloticus) fillets[J]. J Food Prot,2019,82(10):1697−1705. doi: 10.4315/0362-028X.JFP-18-365
[49] 肖咏梅, 李明, 毛璞, 等. 黄酮类化合物生物改性及活性的研究进展[J]. 河南工业大学学报(自然科学版),2019,40(2):123−131,139. [50] 马艺超, 路飞, 马凤鸣, 等. 体外模拟消化对苦荞面包黄酮及抗氧化的影响[J]. 中国粮油学报,2019,34(9):20−27. doi: 10.3969/j.issn.1003-0174.2019.09.005 [51] 蔡亭, 汪丽萍, 刘明, 等. 超微粉碎对苦荞多酚及抗氧化活性的影响研究[J]. 中国粮油学报,2015,30(10):95−99+106. doi: 10.3969/j.issn.1003-0174.2015.10.018 [52] 左蕾蕾, 徐沛, 饶朝龙, 等. 超微粉碎对苦荞米与苦荞壳总黄酮的体外溶出影响[J]. 食品工业,2016,37(6):109−112. [53] 孙丹, 黄士淇, 蔡圣宝. 不同加工方式对苦荞中总酚、总黄酮及抗氧化性的影响[J]. 食品与发酵工业,2016,42(1):141−147. [54] Singh K, Kumar S, Rani A, et al. PHenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H) and catechins (flavan-3-ols) accumulation in tea[J]. Funct Integr Genomics,2009,9(1):125−134. doi: 10.1007/s10142-008-0092-9
[55] 顾继娟. 苦荞籽粒芦丁降解酶的纯化与其酶学性质研究[D]. 咸阳: 西北农林科技大学, 2010. [56] Wang L, Li X, Niu M, et al. Effect of additives on flavonoids, D-chiro-Inositol and trypsin inhibitor during the germination of tartary buckwheat seeds[J]. Journal of Cereal Science,2013,58(2):348−354. doi: 10.1016/j.jcs.2013.07.004
[57] Ren S C, Sun J T. Changes in pHenolic content, phenylalanine ammonia-lyase (PAL) activity, and antioxidant capacity of two buckwheat sprouts in relation to germination[J]. Journal of Functional Foods,2014,7:298−304. doi: 10.1016/j.jff.2014.01.031
[58] 周小理, 方向, 周一鸣, 等. 磁场对苦荞种子萌发过程中黄酮类物质的诱导效应[J]. 食品科学,2012,33(21):20−23. [59] Krahl M, Back W, Zarnk O W M, et al. Determination of optimised malting conditions for the enrichment of rutin, vitexin and orientin in common buckwheat (Fagopyrum esculentum moench)[J]. Journal of the Institute of Brewing,2008,114:294−299. doi: 10.1002/j.2050-0416.2008.tb00772.x
[60] 雒晓鹏, 卜星星, 赵海霞, 等. LED光源对芽期苦荞黄酮合成的影响[J]. 食品科学,2015,36(3):86−89. [61] 吕兵兵, 姚攀锋, 王官凤, 等. 光周期对苦荞芽菜生长与品质的影响[J]. 西北植物学报,2019,39(10):1785−1794. [62] 卞紫秀, 马辉, 汪建飞, 等. 超声结合NaCl处理对苦荞麦萌发及芽苗主要成分的影响[J]. 安徽工程大学学报,2019,34(5):8−18. doi: 10.3969/j.issn.2095-0977.2019.05.002 [63] 马辉, 卞紫秀, 陈雪怡, 等. 微波协同L-pHe处理对萌发苦荞中主要营养成分的影响[J]. 安徽工程大学学报,2019,34(6):1−7. doi: 10.3969/j.issn.2095-0977.2019.06.001 [64] 张采琼, 赵江林, 赵钢, 等. 香菇多糖对苦荞萌发及黄酮合成的影响[J]. 食品工业,2014,35(8):123−126. [65] 卞小稳. 荞麦在啤酒酿造中的应用研究[D]. 无锡: 江南大学, 2016. [66] 张燕莉. 苦荞啤酒浸麦、糖化工艺优化及酿造过程活性成分变化研究[D]. 合肥: 安徽农业大学, 2013. [67] 殷培蕾. 苦荞醪糟发酵工艺及质量评价[D]. 成都: 西华大学, 2015. [68] 杨芙莲, 刘旭. 荞麦醋发酵过程中芦丁含量变化规律研究[J]. 中国酿造,2012,31(9):44−46. doi: 10.3969/j.issn.0254-5071.2012.09.013 [69] 王延丽. 我国荞麦食品的加工研究[J]. 食品工业,2016,37(5):267−269. [70] 马艺超. 不同热加工对苦荞制品功能成分、质构及体外消化的影响[D]. 沈阳: 沈阳农业大学, 2019. [71] Bouasla, A W. Rice-buckwheat gluten-free pasta: Effect of processing parameters on quality characteristics and optimization of Extrusion-cooking process[J]. Foods,2019,8(10):496−496. doi: 10.3390/foods8100496
 下载:
下载:

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
